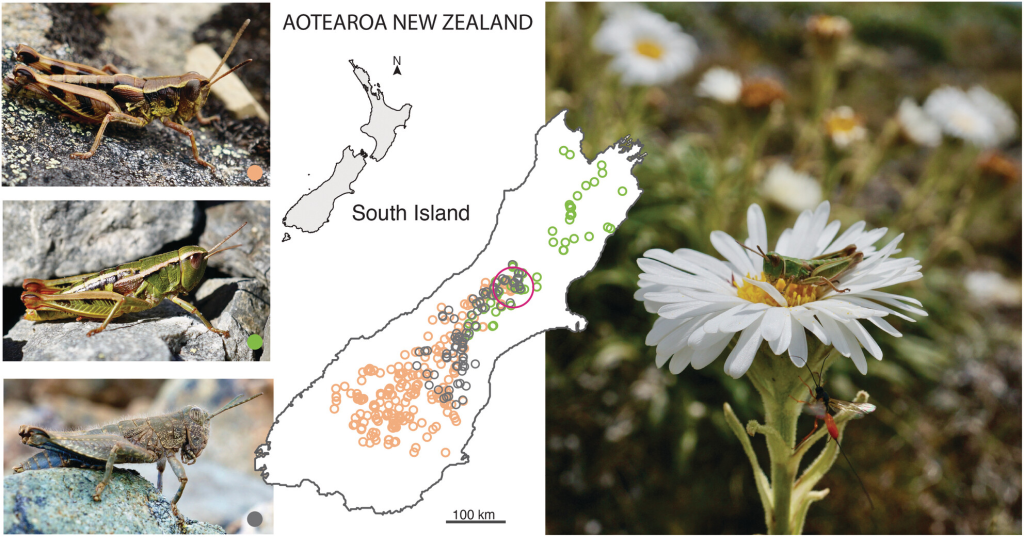
A study of grasshopper diet compared what New Zealand alpine grasshoppers were eating 50 years ago with todays meals. The Massey University biologists found that exotic weeds that were rare or absent in Canterbury mountains 50 years ago are now an important part of the diet of native grasshoppers.
None of the alpine grasshoppers studied liked eating tussock despite it being the most common plant in their environment. The grasshoppers preferred to eat soft weeds such as hawkweed, catsear, sheep sorrel and false dandelion – new plants spreading into New Zealand high country during the last 50 years.
The food eaten by three species of grasshopper was studied using microscopic examination of plant fragments and DNA sequencing. Both methods showed that grasshoppers select soft herbs and avoid the tough tussock and spiky speargrass of the mountains. In the 1970’s the grasshoppers ate lots of native herbs, but these days the same species of grasshoppers are eating more weed species. By eating flowers, fruit and leaves of soft plants the grasshoppers are helping the tussock plants have space to establish – ensuring New Zealand mountains continue to be covered in snow tussock.
Māwhitiwhiti Aotearoa The flightless and silent grasshoppers studied were three species of the endemic genus Sigaus. Thirteen species evolved in New Zealand and live primarily in the alpine zones of Kā Tiritiri o te Moana the Southern Alps. They are under threat from habitat loss due to anthropogenic climate change.
The plants that are new to New Zealand mountains have become major components of the diet of endemic grasshoppers
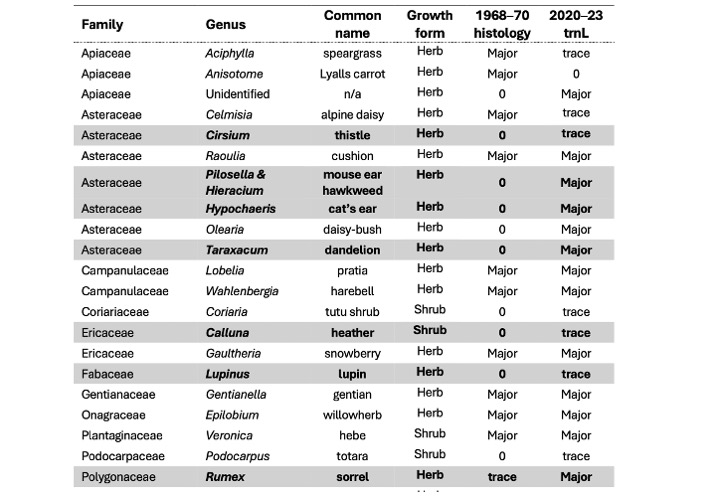
Major components of grasshopper diet recorded from gut content analysis of New Zealand alpine Sigaus grasshoppers using microhistological analysis (1968–1970) and DNA metabarcoding with trnL (2020–2023). Introduced invasive plants species in New Zealand highlighted with grey rows. Components of diet divided into Major, trace or not detected (0).
Ground wētā are those species of flightless crickets endemic to Aotearoa-New Zealand that conceal themselves in daytime in soil burrows. Recently we showed that this lifestyle has been adopted by two separate evolutionary lineages in NZ; Hemiandrus and Anderus.
Though species in these two genera are all considered to be ground wētā, each group is more closely related to separate king crickets in Australia that to each other. The name Hemiandrus was established in 1938 but Anderus was formally recognised more recently in 2024.
Hemiandrus dryadis photographed by Richard Littauer https://inaturalist.nz/observations/259419032
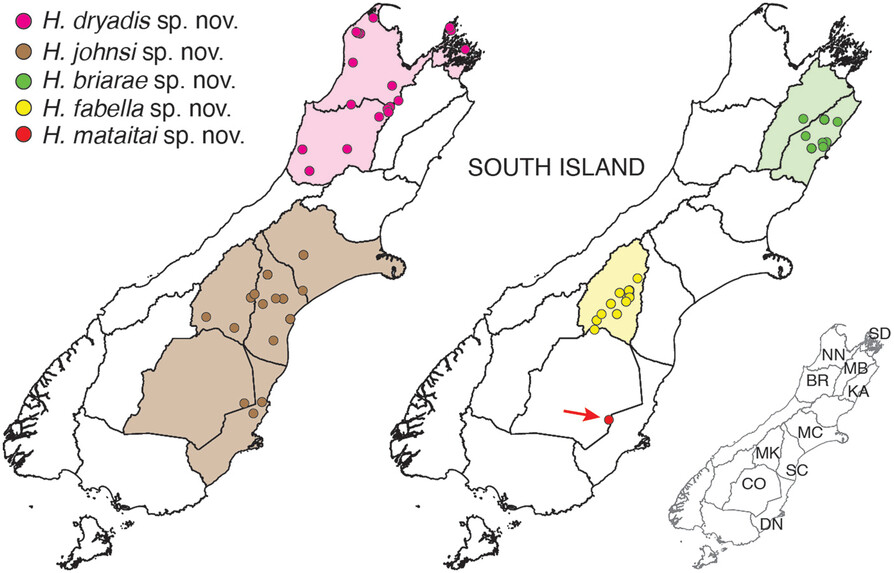
The Hemiandrus group now includes 16 described species with the recent addition of five new names. One of these is the svelte Dryad’s wētā (Hemiandrus dryadis) that lives in native forest habitat in northwest South Island. Orange hued and long-limbed, this species is most similar to Jacinda’s wētā in North Island.
A second large species is Briar’s wētā which is lives in neighbouring regions in northeast South Island, but appears to be limited to high elevation habitat. It lives amongst sub-apline vegetation, which suggests it is cold tolerant and could be freeze-tolerant like other New Zealand alpine insects.
Hemiandrus briarae photographed by Carey Knox https://inaturalist.nz/observations/262763151
Three other smaller species are from low lying areas in Canterbury and Otago and these include a species associated with the Tekapo river area and sometimes referred to as the. ‘Tekapo’ wētā. This species now has the formal name Hemiandrus fabella, which means ‘little bean’ in reference to its small size and compact shape.
The known distribution of five new species of Hemiandrus ground wētā in South Island, New Zealand. Spots are known records, and coloured areas are recognised entomological regions.
All of these species are threatened by human activities that modify their habitats, and predation by introduced pests such as hedgehogs. In particular the lower lying areas of Canterbury region are subject to ongoing agricultural intensification so the current and future status of H. fabella, H. johnsi and H. mataitai remains uncertain. Biodiversity can be lost before being discovered, and the Sutton Salt Lake wētā H. mataitai , which is known from just one location is an example of the fragility of the situation.
REFERENCES Trewick SA, Morgan-Richards M. 2026. Five new species of New Zealand Hemiandrus Ander 1938 ground wētā (Orthoptera: Anostostomatidae). NZ Journal of Zoology 53:e70007. https://doi.org/10.1002/njz2.70007 Trewick SA, Morgan-Richards M. 2025. Two new species of New Zealand Anderus Trewick et al. 2024 ground wētā (Orthoptera: Anostostomatidae). Zootaxa 5666 (3): 408–418. https://mapress.com/zt/article/view/zootaxa.5666.3.5 Trewick SA, Taylor-Smith BL, Morgan-Richards M. 2024. Wētā Aotearoa—Polyphyly of the New Zealand Anostostomatidae (Insecta: Orthoptera). Insects 15: 787. https://www.mdpi.com/2075-4450/15/10/787 Trewick SA. 2021. A new species of large Hemiandrus ground wētā (Orthoptera: Anostostomatidae) from North Island, New Zealand. Zootaxa 4942 (2). https://www.biotaxa.org/Zootaxa/article/view/zootaxa.4942.2.4
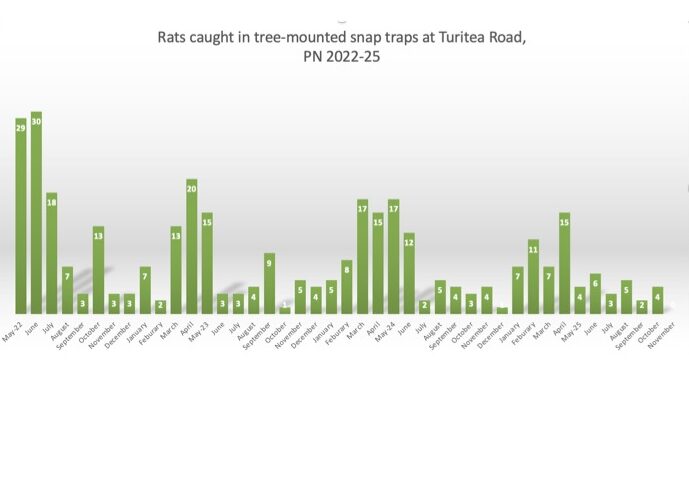
Good news from Turitea Valley to end 2025. November 2025 is the first month since May 2022 (when I began using trunk-mounted snap-traps) that I recorded a glorious zero. No rats! As of Christmas day there have been no rats this month either, but I’m not counting my chickens.
Song thrush eggbird egg after rat has eaten embryo
In October I not only killed four rats in snap-traps but also saw signs of rat damage (broken thrush egg). In the Green Corridors by Turitea Stream there has been steady numbers of rodents killed in stoat traps (DOC200) through spring and early summer. So I know that rodents are nearby and will arrive on my property, but for now I’m celebrating a new record and (hopeful) that there is a lower rat density at my place as tui, grey warbles and pīwakawaka feed their fledglings.
After publishing the species description we realised that we had re-described Celatoblatta undulivitta Walker 1868. We made a mistake – the species that needs a name is the very similar but more widespread in North Island New Zealand. In 2026 we’ll put this right.
The discrepancy between formally described species and the number of species that actually exist is known as the “Linnaean shortfall”. In Aotearoa New Zealand, it is 59 years since the last cockroach species was formally described [Morgan-Richards & Trewick 2025]. Some might think that another cockroach is the last thing we need but taxonomic knowledge of all dimensions of life on Earth is critical to cataloguing, measuring, and understanding biodiversity. Biodiversity exists and documenting it is critical, but taxonomy is a field without funding.
Two irregular dark stripes on the face of the new species simplify its identification
A new species of forest cockroach endemic to Aotearoa New Zealand is more than just one new species we need to save from extinction. This insect (~ 15mm long) is home to its own species-specific Blattabacterium endosymbionts; an ancient heritage of the cockroach-termite lineage dating back more than 200 million years. These bacteria live only inside body tissue of each cockroach and contribute to its survival. In addition, like other animals including humans, each cockroach hosts a novel community of microbes in its gut.
The new kauri cockroach Celatoblatta kaurijust described is almost certainly the host of unique Blattabacterium and other forms of micro-biodiversity. These particular organisms are restricted to northern New Zealand where kauri forests grew before humans arrived. Each time we name a new species we create the words needed to describe the intimate associations between host and microbe.
The practice of coining a Latin binomial to apply to a newly described species has consequences for the science we do, influencing its resourcing, its public communication, and sometimes even the direction of research. The lack of support for taxonomy and the resulting slow rate of species discovery and description is a phenomenon referred to as the ‘taxonomic impediment’. Naming biodiversity should be a priority in conservation biology because until we know what we have we do not know what we are trying to save.
The noble false widow spider (Steatoda nobilis) has found urban habitats profitable and has become one of the most successful invasive spiders around the globe. One that is continuing to expand its range thanks to the transport and habitat given it by humans, and one that carries with it a menagerie of microbes.
Invasiveness combines an ability to get to a place and ability to survive and breed there. Animals that associate closely with humans are referred to as synanthropic and are especially good at travelling with people and so continue to share their urban environment, wherever in the world that is. (Right: Male Steatoda nobilis from Porirua, Wellington. (c) S.A.Trewick).
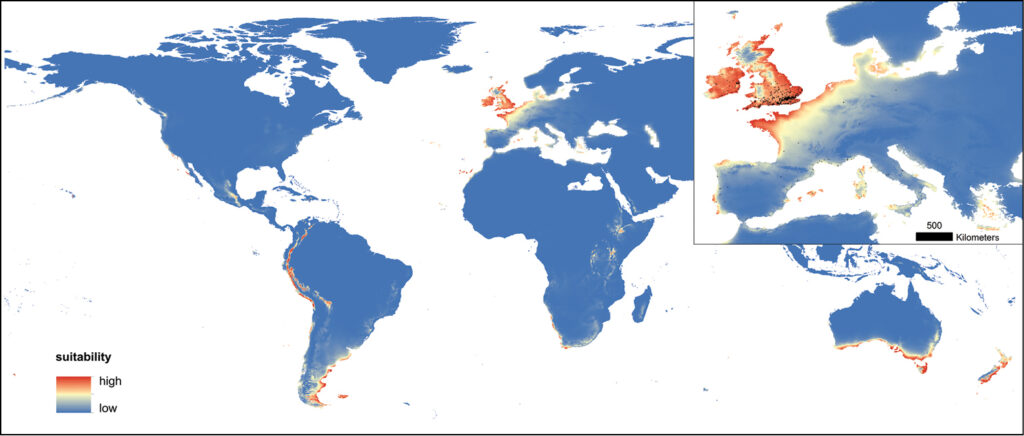
Humans unwittingly enable certain species to invade, but, it appears that human environments are not all equal. Species such as the noble false widow cannot thrive everywhere; they have physiological limits and climatic conditions are especially important for ectothermic creatures including spiders. Planet-scale climate influences where the most suitable habitat is.
Using global climate data and information from from locations where Steatoda nobilis is found it is possible to model where conducive habitat exists around the globe (Bauer et al. 2019).
Remarkably, a study published in 2019 using ecological niche modelling predicted that many parts of New Zealand were likely to be suitable for Steatoda nobilis. They have been proved right; the noble false widow is living in cities across New Zealand, nestled in urban gardens and around houses.
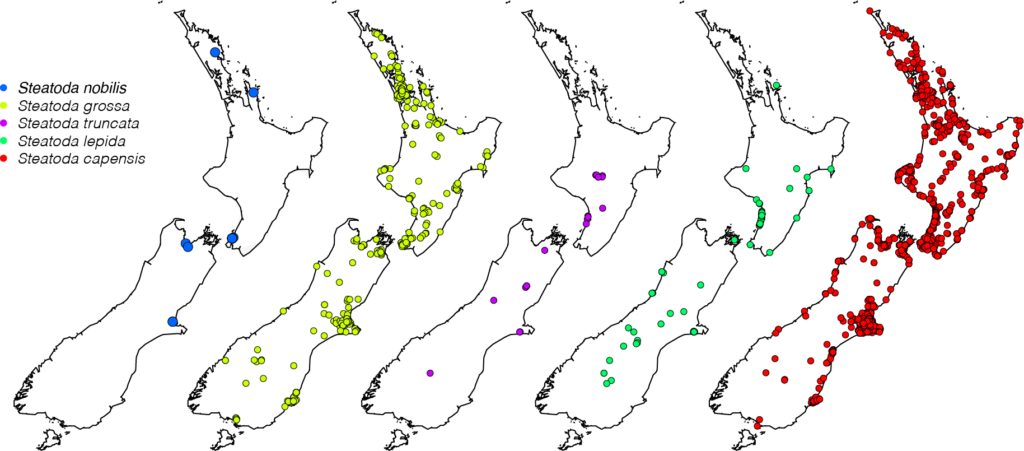
Research quality observations of invasive Steatoda spiders in New Zealand.
New Zealand now has five species of introduced false widow spider, all associated with modified habitats across the country. Each must have arrived by some form of accidental transport, but then colonised successfully.
False widow spiders are so-called because they resemble Latrodectus widow spiders such as the black widow L. lilianae (Left. (cc) Jorozko). The similarity does not end there. Although generally considered less dangerous than widow spiders, the venom of Steatoda nobilis contains toxins similar to those produced by Latrodectus lilianae.
The spread of Steatoda nobilis is paralleled by increasing concern about its human health and ecological impact as an invasive species. Not only can the species inflict a painful bite with Latrodectus-like symptoms but it has emerged that associated bacterial infection can lead to necrosis (tissue death). Of 22 bacterial species detected on S. nobilis, 12 are known to be pathogenic in humans. Sometimes the infections resulting from bites do not respond to standard antibiotic treatments, and the level of antibiotic resistance displayed by some Steatoda-associated bacteria is significant for medical science.
Antibiotic resistance profile of the bacterial community isolated from body and chelicerae of Steatoda nobilis. (A) Number of bacterial isolates resistant to each antibiotic. (B) Number of isolates showing resistance to 0, 1, 2 and 3 diferent antibiotics. Dunbar et al. 2020.
It is also significant for ecosystems. Biodiversity loss and accelerated environmental change are hallmarks of the Anthropocene, and rapid global climate shifts are expected to increase the rate at which invasive species replace native species.
Female Steatoda nobilis from Porirua, Wellington. (c) S.A.Trewick
Brushtail possums (Trichosurus vulpecula) are one of the most troublesome pests in New Zealand and despite the governments ‘PredatorFree2050’ aspirations we are a long way from eradicating this invasive species. The very effective use of synthetic fluoroacetate (1080) poison has short-term impact on total numbers.
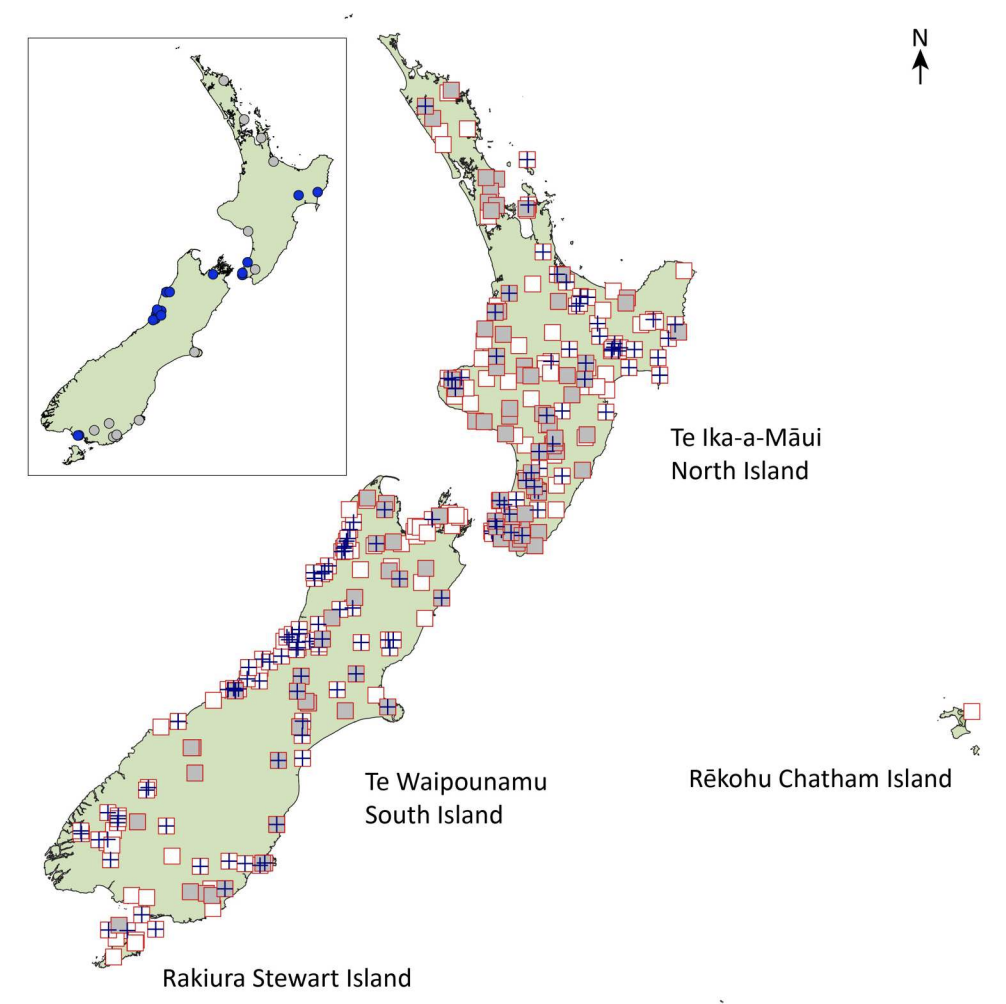
Invasive pests commonly result from accidental introductions or opportunistic colonisation involving few individuals, but Acclimatisation societies worked assiduously to import and release brushtail possums throughout the country.
Documented release sites (red squares) of brushtail possums in New Zealand between 1865 and 1952. Black possums (dark blue cross) and grey possums (grey fill) are indicated where known. Inset map shows locations of first introductions from Tasmanian (dark blue) and Australian (grey) importations made between 1837 and 1898. Pattabiraman et al. 2025
‘We shall be doing a great service to the country in stocking these large areas … with this valuable and harmless animal’ reported the Auckland Acclimatisation Society in 1917. New Zealand forest were considered to be under utilised and a widely held belief was that a valuable fur trade would result from possums. The legacy of this naivety is a species that damages New Zealand agriculture, tourism, biodiversity and ecosystem resilience through transmission of bovine TB, spread of bacterial pollutants to waterways and destruction of native animal and plant populations.
A view of the current success of possums in New Zealand can be found here: PossumNZ
The efforts of the individuals and societies primarily in the latter 19 Century equipped the invasive New Zealand possum population with huge genetic diversity. Multiple origins of the possums in Australia are evident from the variation in fur colours, that reflect different subspecies. Release in New Zealand created a genomic melting pot that enabled mixing of of these variants which freely interbreed. Evidence for this mixing comes from population genetics, and analysis of morphological evidence that show a continuum in variation.
Examples of naturally occurring fur colour variants among invasive brushtail possums in Aotearoa
Fur colour is probably of little consequence to possum survival in New Zealand but it provides a visual clue of genetic diversity. Possibly the most concerning genetically controlled attribute of brushtail possums for New Zealand is in there potential to tolerate high levels of 1080 poison.
So far, the fact that possums came from southeast Australia and Tasmania has benefited control measures as possums in those areas have low tolerance of natural 1080 because it is rare in the plants they eat. The genomic potential for resistance is obvious from possums in Western Australia that eat plants that naturally have high 1080 levels. WA possums have high tolerance of the poison. See POSSUMS STOMACH 1080
Remarkably, since the 1980s no new data have been obtained that directly tell us how tolerant of 1080 possums in New Zealand are. In fact the research done in the development of 1080 baits in New Zealand has never been published and only summary data are available (captured in table above).
Climate change is here. Increasing global temperatures has already resulted in alpine species disappearing from their lowest elevations.
In Aotearoa New Zealand this is most apparent among the endemic alpine-adapted grasshoppers, that live above the tree-line on mountains and have evolved to survive repeated freezing and thawing.
The black-eyed alpine grasshopper Sigaus villosus lives only on the tops of New Zealand mountains.
Data spanning 52 years show the lowest elevation recorded for New Zealand’s largest grasshopper (Sigaus villosus) in Canterbury has moved 290 metres up the mountain. This means this flightless black-eyed alpine specialist is now missing from habitat it was using in the 1960s because rising temperatures have already reduced the habitat suitable for this endemic species.
Although some alpine species can move up mountains to track cooler conditions higher up, that is only possible if the mountain is tall enough. In some places they have reached the top already, resulting in a squeeze on space as lower elevations get too hot. This change to the distribution of alpine insects is documented world-wide as shown by a newly published review in the science journal Evolution & Ecology: Meza-Joya et al. 2025
The review of alpine insect species from around the world shows that more than half have lost some of their lowest elevation habitat (over 10 or more years). Upslope expansion was observed in 56% of studied alpine species – but upslope expansion has not been recorded for New Zealand’s largest alpine grasshopper which can live at 2130 m above sea level. Fewer than 30 peaks in the Southern Alps are over 3000 m asl so there is not a limitless supply of potential habitat.
Space is not the only problem. Higher up a mountain the lower the oxygen level – a factor that might prevent some species expanding. And, the shape of mountains means that habitat patches get smaller, fragmenting and reducing populations size.
Alpine habitat fragmentation with climate heating. Patches of suitable habitat (PINK) become smaller, fewer and further apart under two future climate scenarios for the endemic, flightless, alpine grasshopper Sigaus australis (Koot et al. 2022; Meza-Joya et al. 2023).
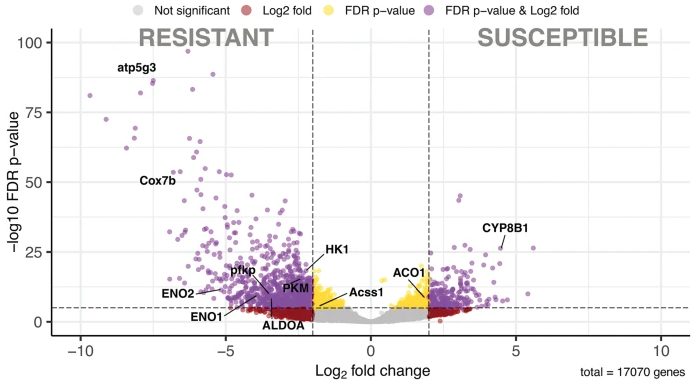
Multiple genes have been identified that might explain why some possums can tolerate high doses of plant toxins. The scientists compared functioning genes from wild brushtail possums (Trichosurus vulpecula) that came from east and west Australia, known to have contrasting tolerance to 1080. The lethal does of 1080 for brushtail possums in WA is about 150 times higher than for eastern possums.
By comparing genomic data from possums in these different populations it was possible to identify genes that are expressed differently. The research focussed on gene expression in the liver which is recognised as a centre of toxin breakdown in mammals. Some of the differences in expression that were found are associated with differences in the response of the possums to what they eat.
A suite genes are expressed more in possums from Western Australia (left) where they have evolved high tolerance (resistance) to naturally Compound 1080 (monofluoroacetate) produced naturally by native plants. (from Carmelet-Rescan et al. )
1080 is a mammal toxin that is naturally produced by many plant species to protect their leaves from leaf-eating animals. It is more abundant in plants in Western Australia than east Australia, where the possums introduced to New Zealand came from.
Animal cells use a series of chemical reactions known as the citric acid cycle to release energy from nutrients including carbohydrate, fat, protein and even alcohol. That means this system is not only the powerhouse of animal life but it contributes to the break-down of a wide range of compounds some of which are toxins. 1080 produced by plants disrupts this critical mechanism. Adaptations by West Australian possums involve genes associated with the citric acid cycle linked to toxin tolerance.
The research indicates a number of different genes linked to the citric acid cycle are involved in the greater tolerance to 1080 displayed by West Australian possums. Although no WA possums were brought to New Zealand, the NZ invasive population is genetically diverse because of the number and range of possums originally introduced from southeast Australia and Tasmania. The resulting genetic mixture means there is lots of variation on which natural selection can act.
Next steps in this research are to measure expression of implicated genes in New Zealand possum populations.
Possums are omnivores that eat native plant and animals that reduce resilience of our ecosystem. They are also the primary carrier of the infectious disease Bovine tuberculosis in New Zealand so a major concern for agriculture and the economy. The main method of control across Aotearoa New Zealand is aerial application of Compound 1080- monofluoroacetate.
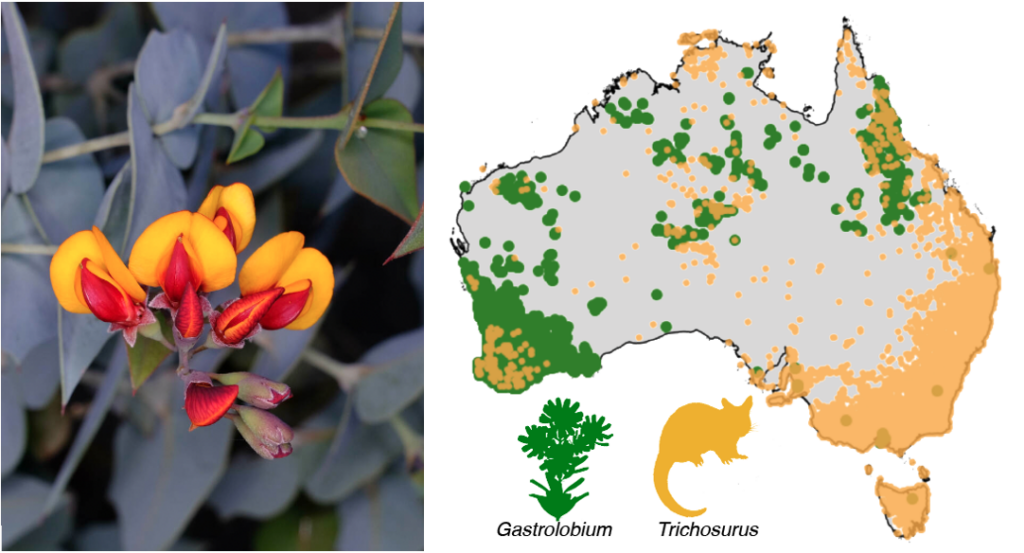
In Western Australia plants such as Gastrolobium spinosum (credit Peter Crowcroft) that belong to the pea family , are toxic to many animals as was discovered in the 1840’s when farm stock died. Native animals in the area are tolerant of the chemical – 1080. Possums (yellow spots) occur across Australia but 1080-bearing plants (green spots) are mostly in the west and north.
Although a source of controversy the Parliamentary Commissioner for the Environment report in 2011 came out in support of continued use of 1080 as the best solution to a major pest problem. Bearing in mind the proposal to eradicate possums and several other introduced mammals from New Zealand by 2050, 1080 remains a priority tool. However, there is the possibility that possums in New Zealand will evolve increasing resistance to 1080 as a result of repeated exposure around the country. Natural selection will favour any individuals with genetics that helps their survival, so it is probable that resistance to 1080 will emerge. Evolutionary responses to human actions are documented in many different circumstances including insecticide resistance in the Lucilia flies that cause fly strike in sheep, and bacterial resistance to most antibiotic medicines.
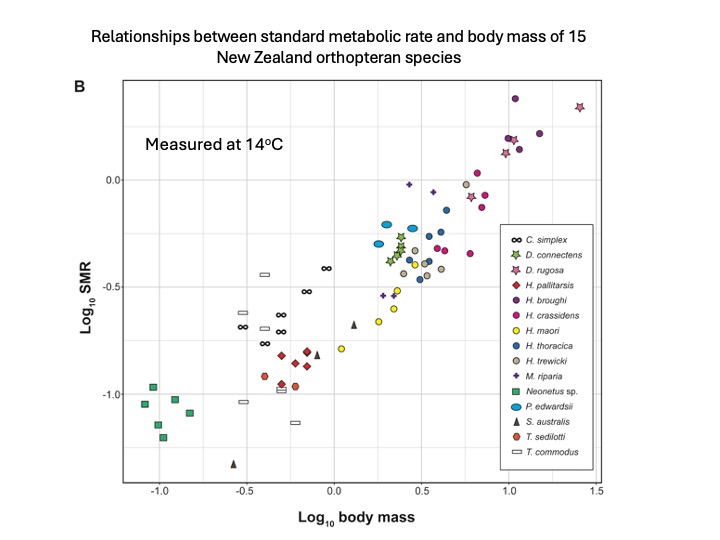
New research shows that New Zealand Orthoptera don’t conform to the metabolic cold adaptation hypothesis.
Metabolic rate varies with body size but when size is considered in concert with other factors many cold-adapted organisms show faster metabolic rate than their warm-adapted cousins at the same test temperature. But this is not what we see in Aotearoa NZ.
In NZ most of our native wētā and cricket diversity is nocturnal and many species are tolerant of cool temperatures or even thrive in cold places. However, their metabolic rates do not support the theory that to cope with the challenges of the cold, alpine insects run their engines faster.
Grasshopper, cricket and wētā energy expenditure is size dependent
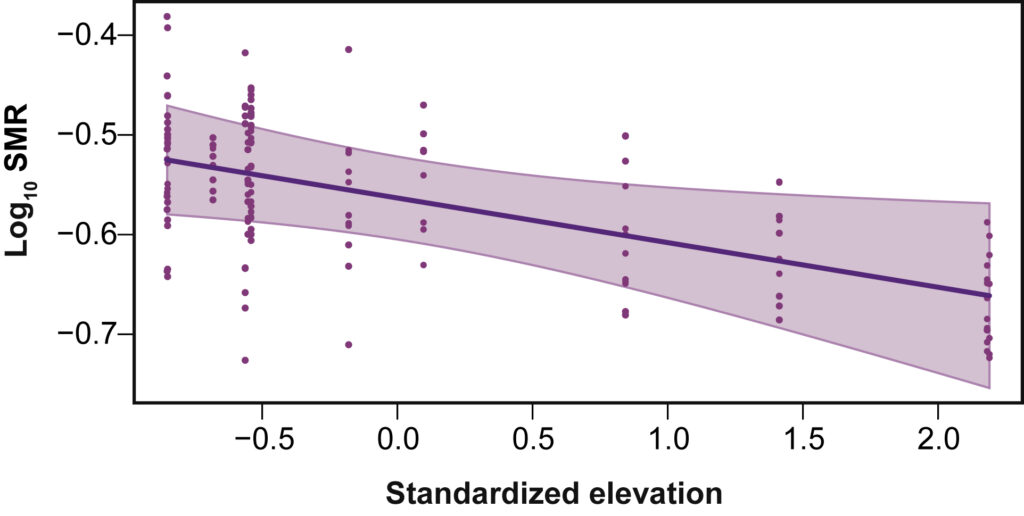
The metabolic cold adaptation hypothesis predicts that ectotherms from colder climates (high latitudes or elevations) have steeper thermal performance curves and elevated metabolic rates at the same test temperature compared to those from warmer environments . We found that New Zealand insects from higher elevations and latitudes had lower standard metabolic rates than expected.
Alpine Orthoptera in NZ have lower metabolic rate than their lowland relatives
As our planet rapidly warms we need to consider the thermal sensitivity of our wildlife. This is the capacity of an individual animal to change its metabolic rate as a response to increases in temperature. Two localized, declining wētā species (Deinacrida rugosa and Motuweta riparia) have high thermal sensitivity of metabolic rate. Climate change will elevate the average temperature wēta experience resulting in increased metabolic rates in these thermally sensitive species, thus requiring greater energy expenditure. This will be an added challenge to species already facing threats from habitat loss and novel predators.
Cook strait giant wētā Deinacrida rugosa is at risk of extinction. Increasing global temperatures will require greater energy expenditure by this insect due to its high thermal sensitivity of metabolic rate
If you read that I had killed 300 rats in two years you might have wondered what I did with all those rodent bodies? The next three good reasons for killing rodents all involve native vertebrates: kahu, ruru and tuna. Fortunately, I killed all 300 rats without using poison so I can feed them to other animals without worrying about accumulation of toxic compounds in the food web.
Reason I kill rodents number four:
After photographing the freshly killed rat I put the carcass out on the hillside (white belly upwards) and the local kahu (Australian swamp harrier, Circus approximans) swoop down and grab a tasty snack.
Kahu gets the dead rats
Reason I kill rodents number five
The bodies of mice and small rattus rattus are left out on the top of a wall in the garden and a couple of ruru (morepork, owl, Ninox novaeseelandiae) stop by early in the evening and take the mammal remains. The ruru first came to our window to catch moths and in the spring they still come for the pururi moths which make a great meal for the little bird.
Ruru gets the mice
Reason I kill rodents number six
Some of the rats I take photos of and ear clipping from are caught in stoat traps in the green corridor. Near where I live the council have fenced off of strip of land beside the stream and planted native trees. The green corridor makes a great place for walking dogs or stretching your legs. The council have included lots of stoat traps in these reserves which catch any mammal small enough to squeeze into the ‘stoat-sized’ hole. So more rats, mice and hedgehogs are caught than mustelids. A few of the rats I remove from the DOC200 stoat traps I throw into the stream for the eels to eat. The native Longfin eel (tuna, ōrea, Anguilla dieffenbachia) is quite common in the local stream
Longfin eel (tuna, ōrea, Anguilla dieffenbachia) get a few rats from council stoat-traps