There were no rodents in Aotearoa/New Zealand before humans arrived. The first people came with Rattus exulans (kiore, pacific rat), then later, different people brought different rodents (Rattus rattus; Rattus norvegicus; Mus musculus/domestics). Now there are rats in every corner of the country.
About two years ago I got some new rat-traps and set them up around my home – near the house and in the forest fragment where I live. I own 2.3 hectares of land that was once used for sheep farming, but the steep river terrace and gully would never have been very productive. The trees and punga in the gully were allowed to grow and they are expanding and replacing the gorse. I caught 29 rats in the first month. I use snap traps mounted on tree trunks and baited with peanut-butter. After two years I have caught 300 rats. All but one of these 300 were Rattus rattus – the ship rat, the black rat, the roof rat, the long tailed rodent. The single Rattus norvegicus was killed in a stoat trap on the ground.

Reasons I kill rats, number one:
A dead rat can’t eat a native insect. In New Zealand forests Rattus rattus spends about 70% of its time off the ground and eats insects and plants. The insects it can find crawling about in the trees at night are tree wētā and ground wētā and cave wētā and stick insects and cockroaches and beetles and moths and caterpillars and crane flies and cicada and lots more. Rattus rattus also eat seeds and fruit and flowers and eggs and little birds in nests. But mostly it eats our native forest ectotherms – unless it’s dead. If the insects survive tonight, they might be food for the grey warbler or the piwakawaka (fantail) or the pōpokotea (whitehead) or tui tomorrow – but these vertebrates are native to New Zealand and hunt during the day. At night the nocturnal insects might be eaten by ruru (owl) or a spider or a gecko. I’m not anti-predation – but I’d rather it was an endemic species who benefited.
Reasons I kill rats, number two:
A live rat is home to the rat flea (Xenopsylla cheopis) and the flea is home to a bacterium called Rickettsia typhi. This bacterium causes the disease murine typhus when infecting humans. Symptoms are fever, nausea, headaches, and muscle pain. Although murine typhus can be fatal if not treated with antibiotics most recover fully. Killing a rat results in the death of the fleas and the bacteria. I don’t know how many fleas on my rats are infected with Rickettsia typhi but disease reduction could save health care costs.
Reasons I kill rats, number three:
Every rat exhales carbon dioxide. 300 rats (about 30kg in total) are no longer contributing to greenhouse gas emissions. The invertebrates that the rats would have eaten get to walk away – they also release carbon dioxide but they are ectotherms, so their metabolic rate is much lower. The fruit and seeds the rats would have eaten might germinate and produce seedlings. By reducing the number of rats in my gully I’m reducing my carbon footprint.

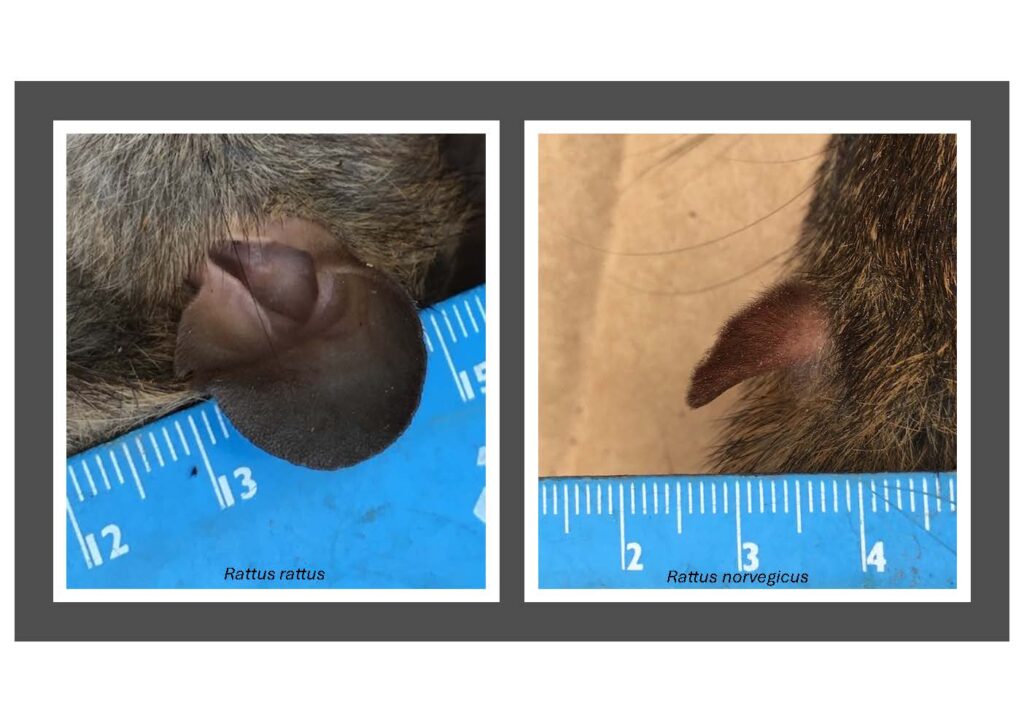
Over the two years I have learnt how to rapidly distinguishing Rattus rattus from Rattus norvegicus – Even if a rat is half grown the tail-to-body ratio, fur colour on belly, shape of face and size of ears will provide the information to separate the two Rattus species.
My two local Rattus species
Although I have killed 300 rats there are still more out there. I know I am not eradicating them from my property as there will always be more moving in. Most Rattus rattus don’t move much further than 100m from their home, if there is plenty of food. But on all sides of my place there are rat populations producing more offspring than can be supported by local food – so from all around me hungry rats will be arriving. The entire rat population was removed from a small Palmerston North forest fragment in 1977 but it took only two months for rats to recolonised the forest (Innes & Skipworth 1983). The graph of number of rats killed per month shows that my local population is responding to the seasons – there is an increase in numbers in early autumn, fewer in winter but never a complete absence. Only if I built a fence could I make a forest fragment free of carnivorous mammals. So, killing rats will not increase the local tui, kereru and the tomtit population but I have three good reasons to bait my traps again tomorrow.

Let the insects live, prevent disease and reduce carbon dioxide:
saving the planet one rat at a time.