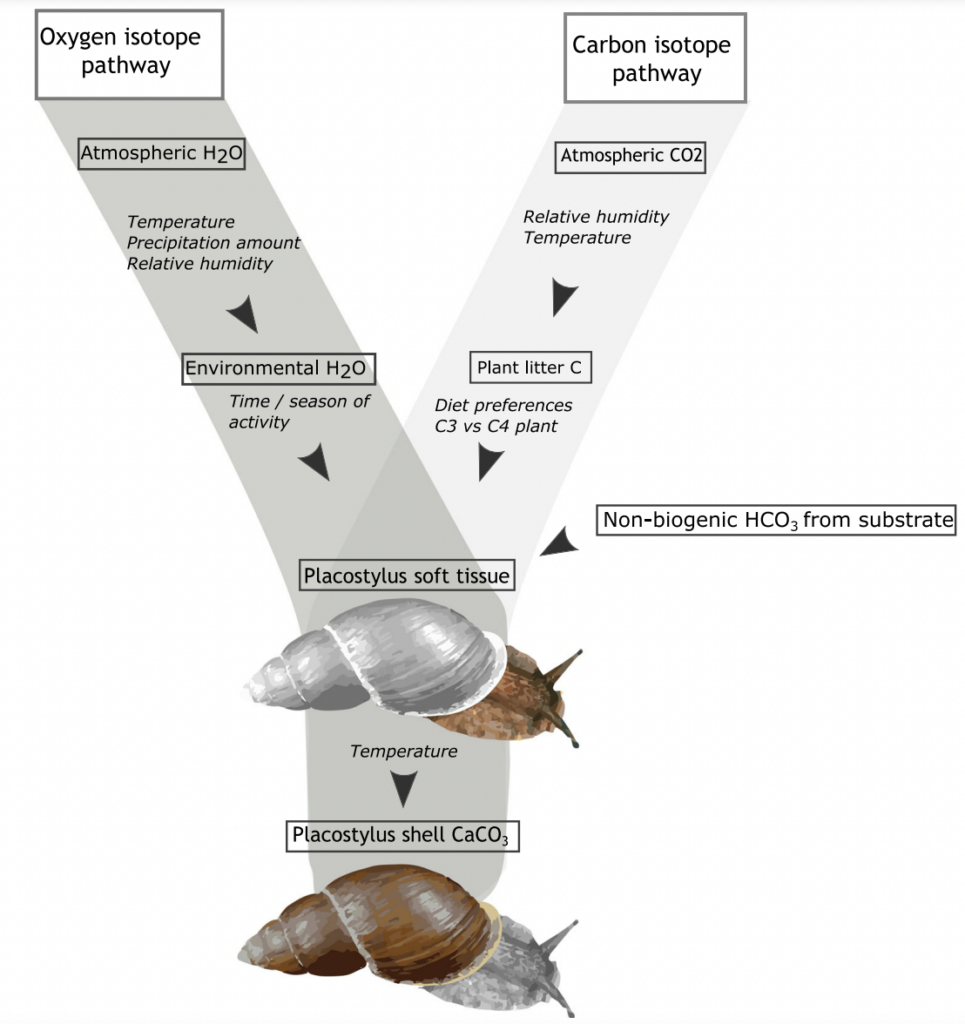
Fossil snails might tell us of the frequency of heavy rainfall in the past
Anew study comparing the stable oxygen and carbon isotope ratios of giant land snails in New Zealand and New Caledonia found a surprising result. New Zealand snails had, on average, higher oxygen isotope ratios values than their counterparts in New Caledonia, counter to the relative isotopic composition of rainwater between these two regions. This research just published in the Journal of Quaternary Science provides baseline data for using the shells of Placostylus snails as environmental proxies – allowing us to use fossil shells to estimate the temperature and rainfall when the snails were alive.
High‐resolution stable isotope profiles from shells of the land snail Placostylus reveal contrasting patterns between snails originating from New Zealand and New Caledonia
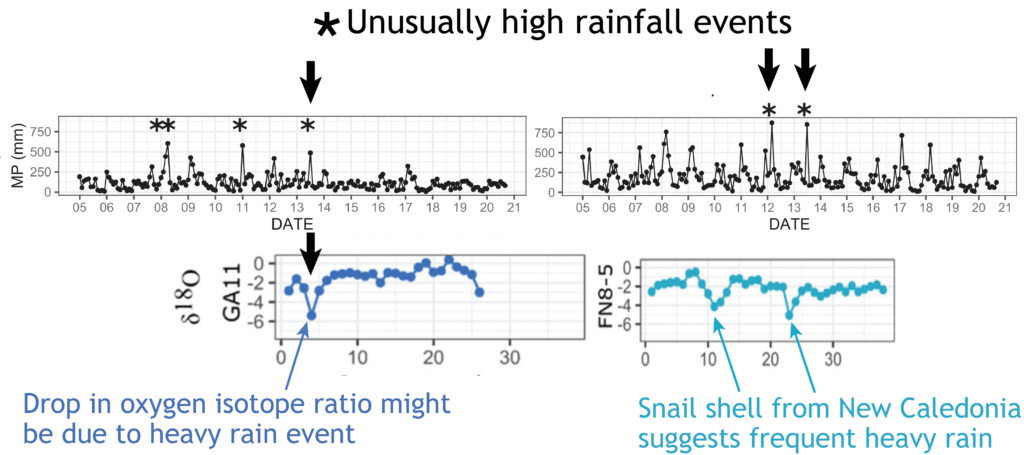
Most interesting are the dramatic drops in oxygen isotope ratios that seem to correspond to heavy rainfall – suggesting an opportunity to dig into the past to compare past precipitation with current frequency of wet weather events in New Zealand. There is also the potential to study the frequency of droughts from the pattern of snail shell growth.
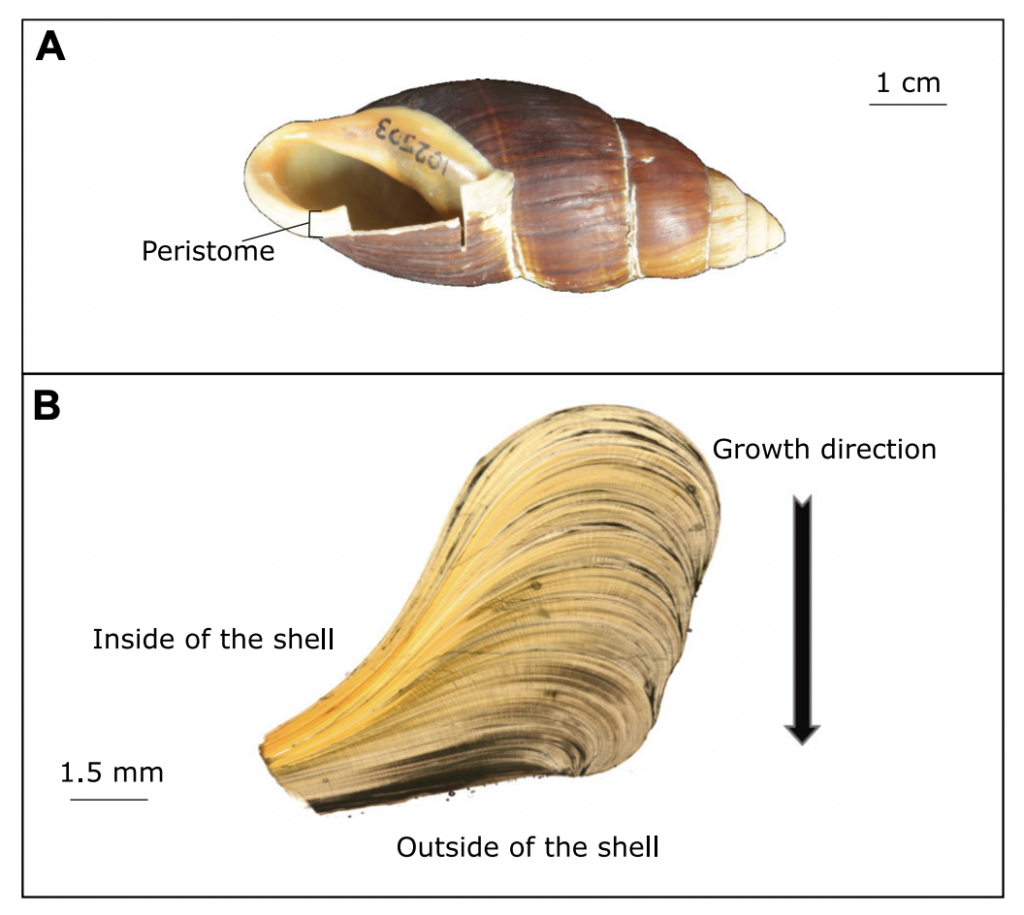
Scientists at Massey University and NIWA sliced up shells of three species of giant land snail (pūpū whakarongotaua; Placostylus). The recent samples from New Caledonia showed drops in isotopic values in their high‐resolution profiles probably linked to periods of intense rainfall.
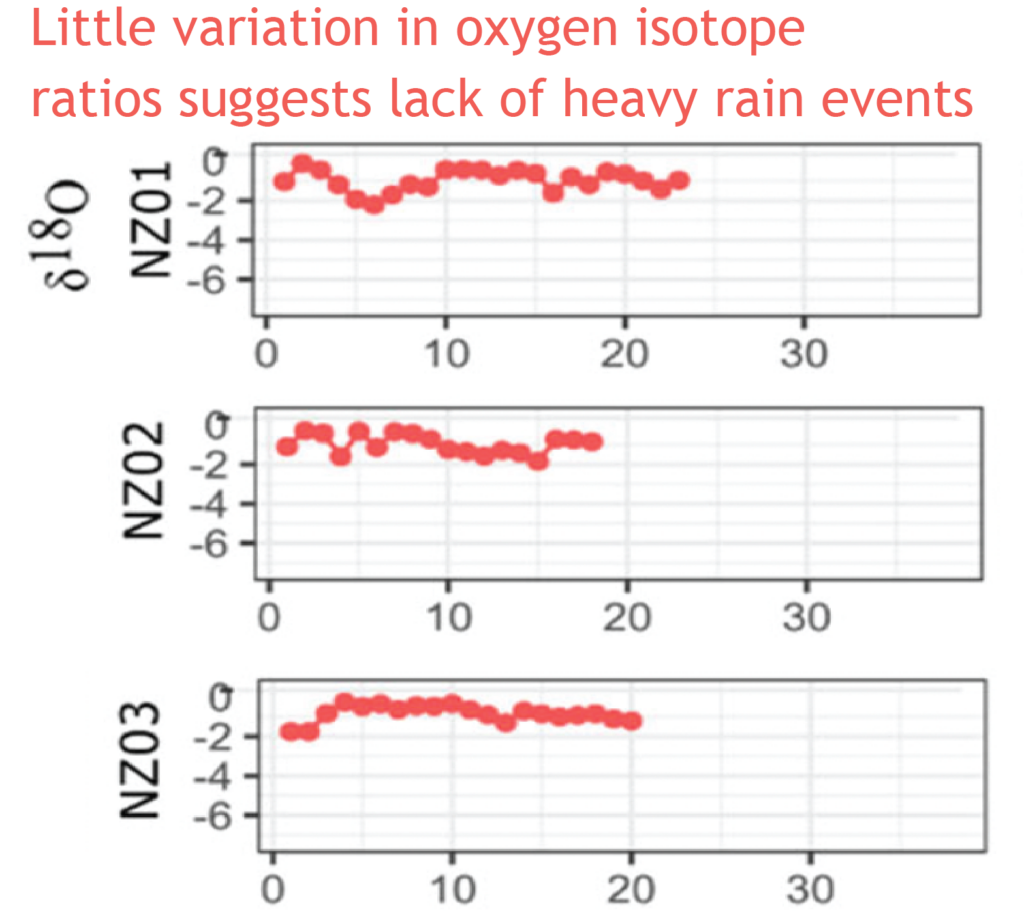
Very heavy rainfall events produce lower stable oxygen isotope ratios incorporated into the shells of the living-growing land snails. In contrast, the snails from New Zealand varied very little, suggesting that when they were alive, 74 years ago, there were few heavy rain events in the Far North of New Zealand.
The snails (pūpū whakarongotaua; Placostylus) are taonga of Ngāti Kurī who value them as security alarms (the snail that listens for war parties). Ngāti Kurī are working to save the local species from extinction but they are also kaitiaki (guardians) of fossil shells buried in the sand dunes and stored in museums. These fossil deposits could provide information about the past climate through high‐resolution stable oxygen isotope profiles.
“it is exciting to think of all the information locked up in snail shells – the shape of the shell, the DNA and the isotopes can all tell a story about the past” said Mary Morgan-Richards. “As Placostylus snails are slow growing, taking 10 years to reach maturity, and live for a long time, they can each tell their own story. There is much to be learnt by digging into old shells to reveal the frequency of heavy rainfall events in the past.”
Global climate change is changing everything, but the gradual processes make it hard to spot the extent of the impact. Change in local climate and the effects it has on species and ecosystems are most apparent where there is a steep gradient in conditions.
A good example of a steep environmental gradient can be found on any tidal rocky coastline, where the tide means some creatures live under sea water, others are exposed to air occasionally, while others that live further up the beach are exposed to drying for much longer. All parts are physically near each other making the gradient in conditions steep. Coastal environments are impacted by global climate change because warming results in the melting of glaciers and polar ice, which in turn leads to rising sea level.
Tree-line on Mount Arthur, Nelson Lakes New Zealand.
On land, a similar situation exists on mountains because the slope of the mountainside means nearby places can have very different conditions. Most obvious as you move up a mountain is the lowering temperature. It is colder near the top than the bottom which is why you go up a mountain to find a ski field.
Another feature of mountains that has a strong influence on biological diversity is their tendency to form ‘islands’; patches of alpine habitat in a sea of lower elevation conditions which in New Zealand is normally forest. Valleys, rivers and forest create a patchwork of mountain tops and ridges; connectedness of these habitats and depends on the climate gradient.
Animals, plants, microbes and fungi that live in the alpine zone, such as this grasshopper (Sigaus piliferus on Mount Ruapehu) have evolved to thrive in the conditions of extreme day-night and seasonal fluctuations in temperature and water availability. Survival means coping with all the different conditions, including being able to freeze when it is cold and re-animate when it is warmer.
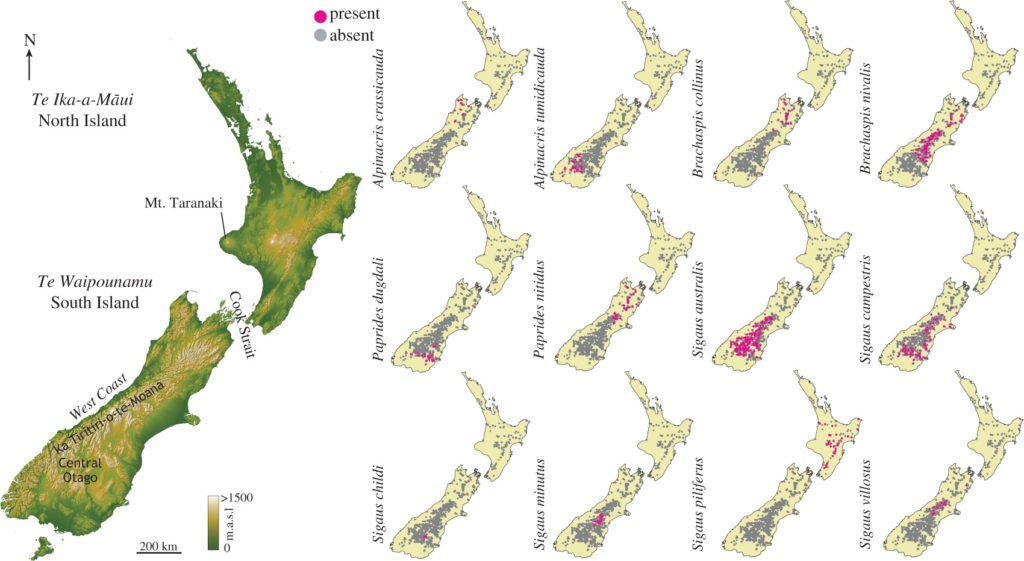
Locations of presence and absence for each of 12 New Zealand grasshopper species
Using the known distribution of New Zealand’s endemic alpine grasshopper species we identified their preferred habitat based on climatic conditions, and then modelled the future status of those habitats given anthropogenic climate warming. We found that available habitat will change for these alpine creatures very quickly; in about 70 years some species could be without suitable habitat that they can use.
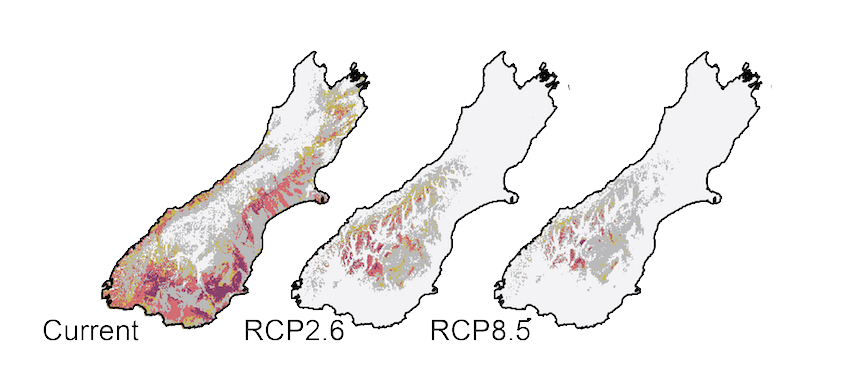
It is easy to measure environmental conditions such as average temperature today, and good data about conditions in the past come fromice-cores and other sources, the future is more uncertain. We know the physics that connects atmospheric gases with global temperature, but the future depends on what people do. We can predict temperature changes during the rest of this century forseveral scenarios which are based upon the balance between the Earth’s heat (energy) gain and its loss (radiative forcing). The most extreme scenario used by the IPCC, RCP8.5, would result with from continued increase in GreenHouse Gases emissions. A more optimistic, but highly unlikely scenario given current trends, RCP2.6, would involve reduction in GHG emissions starting in 2010. RCP2.6 will still result in a 2˚ mean global temperature rise by 2100 (compared to 1750), compared to about 5˚ under RCP8.5.
Current and predicted available habitat for the endemic New Zealand grasshopper Sigaus australis (dark colours most suitable). The RCPB.5 climate change scenario assumes GHG emissions continue to rise through the 21st century. Even under the optimistic RCP2.6 that assumes the C02 emissions started declining in 2020 (they did not) and continue to O by 2100, habitat for S. australis will be scarce. RCP2.6 would result in a global average temperature increase of about 2˚ in the next 70 years.
Our findings apply to all biology living in the alpine zone and, by extrapolation, to all biology in New Zealand. Within one human lifetime, habitat availability will have changed catastrophically for many species… Others may gain, but these will often be species that humans have moved from their native habitat, and become weeds and pests.
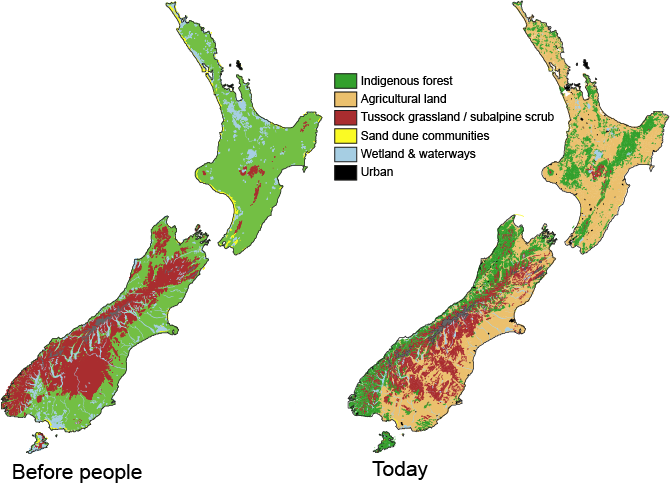
Stand at the side of a mainland island reserve and the impact of humans on New Zealand’s natural environment is obvious. From a landscape naturally dominated by tall forest, agricultural ‘improvement’ rapidly moved us to a uniform, virtual biological desert. Not only are the trees and birds missing, but the lichens, fungi, insects, worms and molluscs are gone. even the bacteria and other microbes of the soil are replaced. In response we resort to counting species and prioritising conservation efforts on the scarcest and restoration effort on the rarest habitats. But, wholesale environmental changes alter no just the abundance of native species but their ecology and interactions. Ultimately, by restructuring the landscape we alter evolutionary outcomes and this has become increasing apparent as research explores biological responses to human induced climate change.
Predator fence on boundary of native forest and exotic paddock. Bushy Park sanctuary near Wanganui.
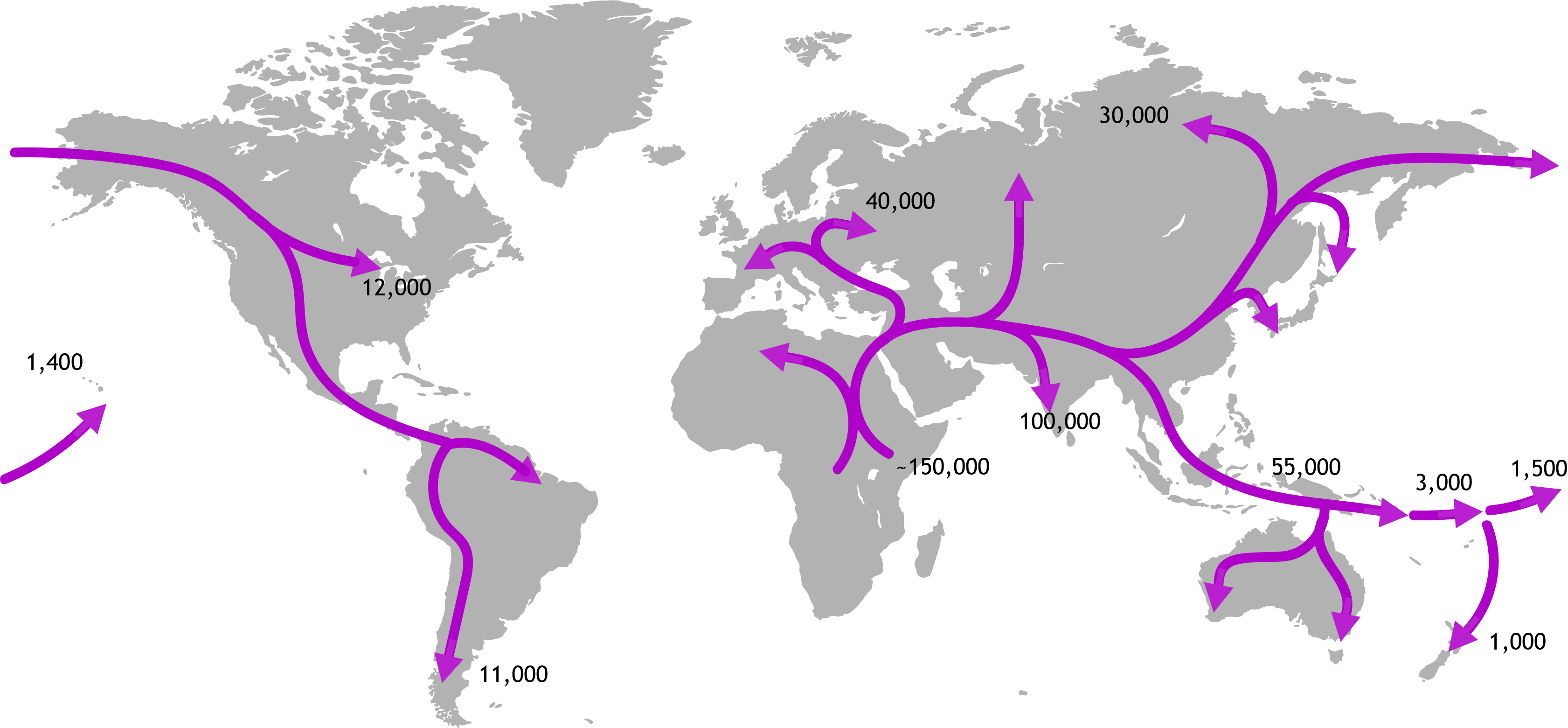
An obvious difficulty with understanding environmental change is that it is much easier to say what is, compared to what was. We are readily inured to the situation and so are accepting of the status quo. One very powerful tool that has helped biologists understand how the geographic ranges of species and population change over time is phylogeography. Simply put, this approach combines information about where individuals and populations of a species are found with information about how those individuals are related to each other. DNA sequence data reveals how closely related individuals are (their genealogy), and how genetically diverse populations are. It is this type of data that shows, for instance, how our human ancestors left Africa and migrated into Europe, then Asia before eventually colonising islands in Oceania. We now know that New Zealand was probably the last major island to have be reached by people travelling by foot and finally boat.
Genetic data from living people has revealed how their ancestors migrated from Africa around the world (values are years before present).
Since the 1990’s phylogeographic studies have revealed the influence of many environmental factors on the distribution of biodiversity. In particular, natural, global climate cycling during the last few million years of Earth’s geophysical prehistory (the Pleistocene epoch) is known to have been influential. We now know for example that in the northern hemisphere repeated extension of the arctic ice cap during ‘glacial’ episodes extinguished populations of all species in northern Europe, Asia and America; remnant populations survived in warmer southern areas. As climate alternately warmed and cooled over 10–100 thousand year cycles, the ranges of animal and plant species expanded and retracted in response.
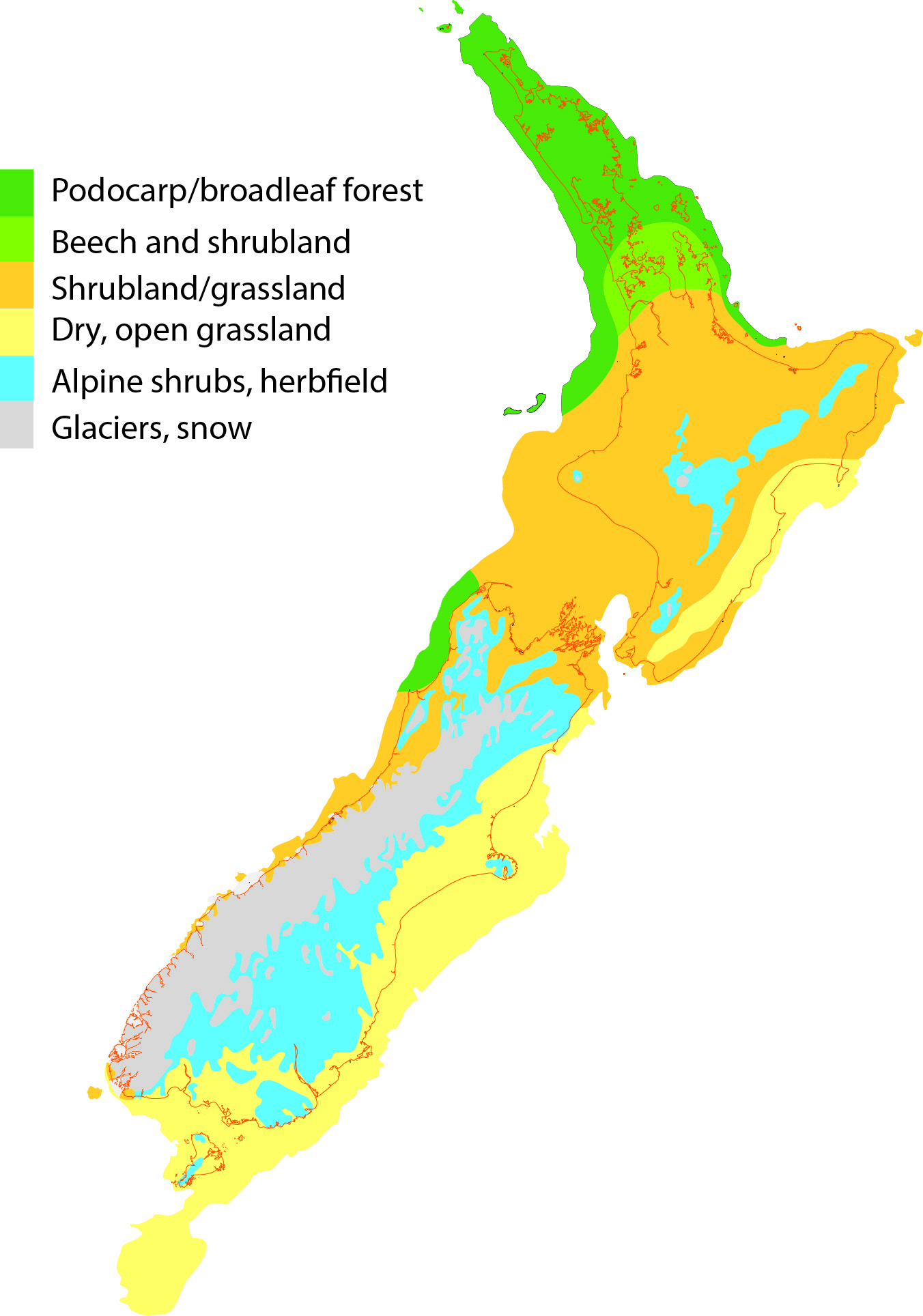
Estimated distribution of vegetation types in New Zealand during the Last Glacial Maximum. See Wild Life New Zealand.
In New Zealand a related pattern of species range change has been inferred. Pollen records show where plant species once lived and genetic data show that during cold phases of the Pleistocene, forest reduced and was replaced in many areas by scrub / grassland communities. Animal species are expected to have responded to these changes tracking their preferred habitat in space and time (or going extinct), and this has been found to be the case for some. North Island tree wētā, for instance, appear to have tracked climate niche.
Phaulacridium marginale
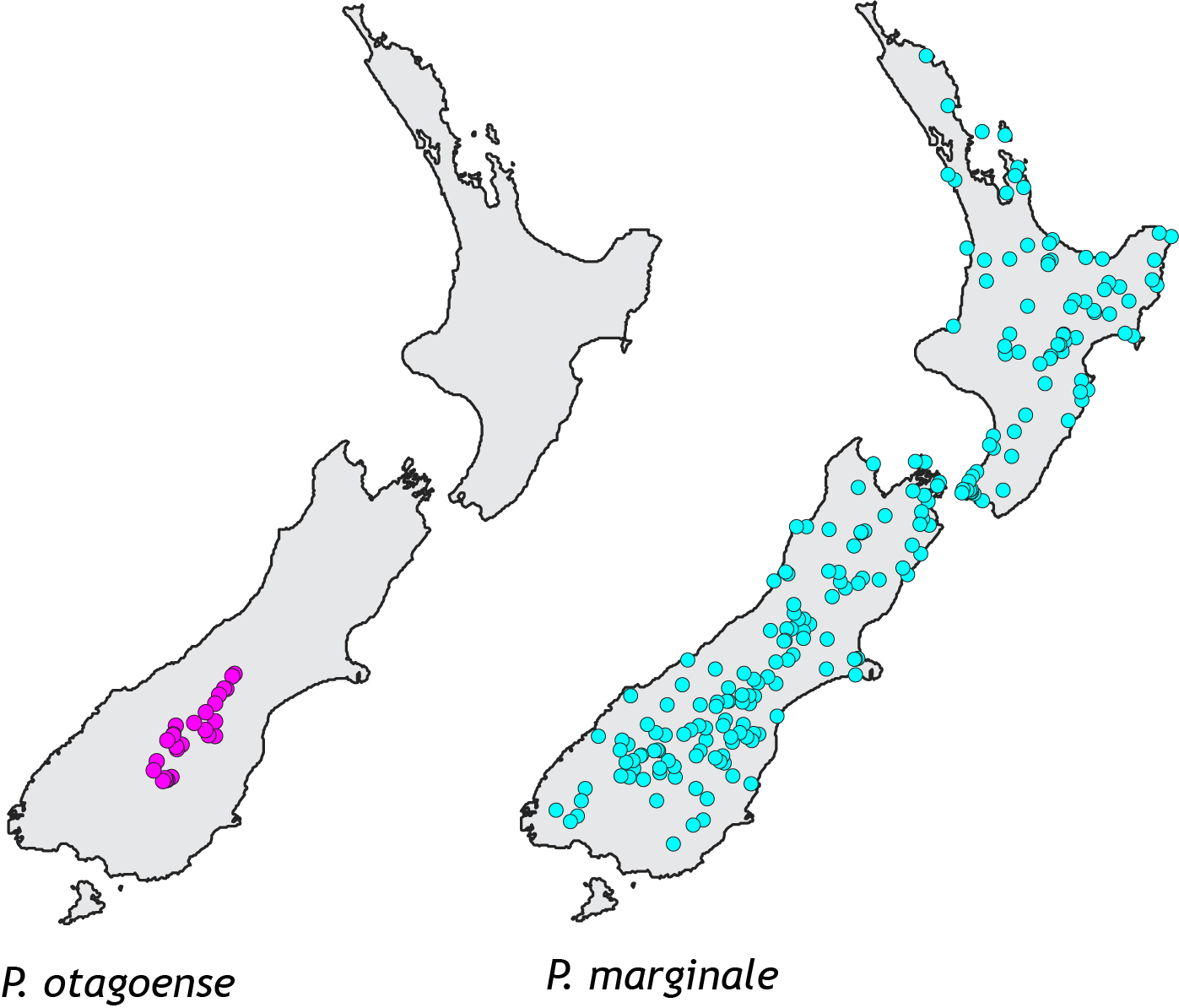
A recent study examined the response of two related grasshopper species. These endemic Phaulacridium grasshoppers live in low elevation habitat, but as is typical of short-horn grasshoppers in temperate regions they require open habitat so they can gain heat by basking in the sun. That means Phaulacridium grasshoppers do not live in forest, and they do not survive above the treeline in the subalpine zone where cool temperatures prevent trees growing (other grasshoppers are adapted to those conditions). So space for Phaulacridium would have been restricted in prehuman New Zealand to scarce open areas such as coastal dunes, river flats, wetlands and semi-arid areas. In fact, one species (Phaulacridium otagoense) occurs today only in the semi-arid McKenzie – Alexandra area of Central Canterbury and Otago. The other species (Phaulacridium marginale) is today found in many places around the country.
Vegetation types across New Zealand before arrival of people (left) and in modern times (right).
Known occurrences of the two New Zealand Phaulacridium grasshoppers.
A small species range usually means a small population size, compared to a species with a big range; and small populations usually have a lower level of genetic variation. Low genetic diversity is documented in many endangered species such as the famous black robins of the Chatham Islands. Paradoxically, in Phaulacridium the opposite pattern exists; the species with the smallest range (pink in map) has much higher genetic diversity than the widespread more common species. The simplest explanation is that P. otagoense (pink), had until recently a much larger range and so bigger population. Conversely, P. marginale (turquoise) appears to have expanded its range recently and has not yet had time to accumulate new genetic diversity.
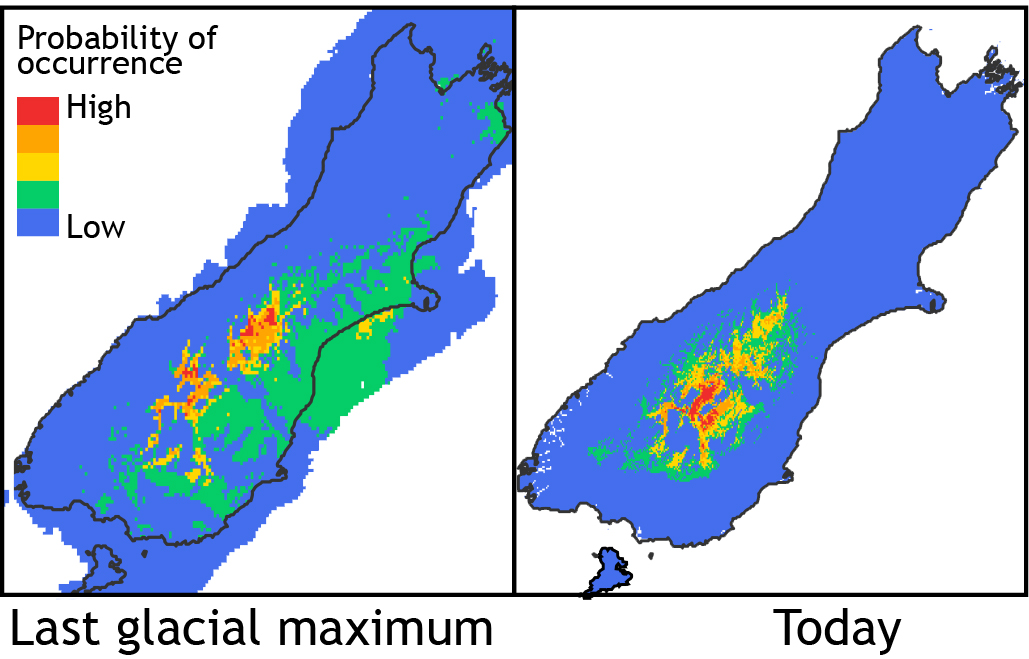
Niche models for Phaulacridium otagoense indicating optimal habitat (red, orange) during the last glacial phase may have been similar to today.
It is known that global temperatures had recovered from the last cold phase of the Pleistocene by about 15,000 years ago. Perhaps P. otagoense had a much larger range in the period before that when cooler, drier conditions allowed scrub grassland to expand; similar to conditions where it occurs today? Niche modelling indicates that in current conditions the potential range of this species is bigger than the actual range in which it is found, and taking into account estimated temperatures during the last glaciation suggests that the habitat preferred by this species had not been much more extensive.
So, probably the major change in fortunes for these Phaulacridium species relates mostly to the recent expansion of P. marginale. Climate modelling shows that the range of this species today is close to the potential occupiable range, but there is a problem. Although the climate across much of New Zealand suits this grasshopper, other factors in the environment do not. In particular, the presence of native forest excludes these little grasshoppers because they need to bask in the sun every day to warm up. How has P. marginale become so abundant and widespread?
The answer lies not in global climate change, but in recent anthropogenic changes to the environment much closer to home. By removing New Zealand native forest, humans created a landscape with the climatic conditions to allow P. marginale to increase in abundance and expand its range across the country. The addition of a mix of northern hemisphere grasses and herbs that thrive in this artificially open environment provided the nutrient-rich food for P. marginale. So that’s great! Well no.
Males and females of different species are capable of reproduction when they meet due to anthropogenic habitat change, resulting in loss of diversity. Male Phaulacridium otagoense with female P. marginale.
The increase in available habitat has meant that the spatial range of P. marginale now meets the range of P. otagoense. Where they meet, the grasshoppers makes mistakes when choosing mates resulting in gene flow. Genetic evidence shows that pure P. otagoense remain in only part of their natural ecological range. Genetic mixing is part of the natural evolutionary mill, but around the world human activity has accelerated the rate at which species meet and interact in new ways. This adds to the trends of biological homogenisation that characterises biodiversity loss in the Anthropocene.