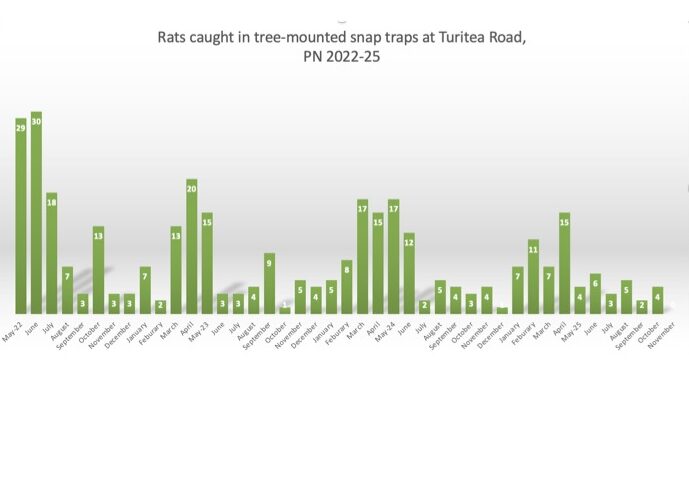
Good news from Turitea Valley to end 2025. November 2025 is the first month since May 2022 (when I began using trunk-mounted snap-traps) that I recorded a glorious zero. No rats! As of Christmas day there have been no rats this month either, but I’m not counting my chickens.
Song thrush eggbird egg after rat has eaten embryo
In October I not only killed four rats in snap-traps but also saw signs of rat damage (broken thrush egg). In the Green Corridors by Turitea Stream there has been steady numbers of rodents killed in stoat traps (DOC200) through spring and early summer. So I know that rodents are nearby and will arrive on my property, but for now I’m celebrating a new record and (hopeful) that there is a lower rat density at my place as tui, grey warbles and pīwakawaka feed their fledglings.
If you read that I had killed 300 rats in two years you might have wondered what I did with all those rodent bodies? The next three good reasons for killing rodents all involve native vertebrates: kahu, ruru and tuna. Fortunately, I killed all 300 rats without using poison so I can feed them to other animals without worrying about accumulation of toxic compounds in the food web.
Reason I kill rodents number four:
After photographing the freshly killed rat I put the carcass out on the hillside (white belly upwards) and the local kahu (Australian swamp harrier, Circus approximans) swoop down and grab a tasty snack.
Kahu gets the dead rats
Reason I kill rodents number five
The bodies of mice and small rattus rattus are left out on the top of a wall in the garden and a couple of ruru (morepork, owl, Ninox novaeseelandiae) stop by early in the evening and take the mammal remains. The ruru first came to our window to catch moths and in the spring they still come for the pururi moths which make a great meal for the little bird.
Ruru gets the mice
Reason I kill rodents number six
Some of the rats I take photos of and ear clipping from are caught in stoat traps in the green corridor. Near where I live the council have fenced off of strip of land beside the stream and planted native trees. The green corridor makes a great place for walking dogs or stretching your legs. The council have included lots of stoat traps in these reserves which catch any mammal small enough to squeeze into the ‘stoat-sized’ hole. So more rats, mice and hedgehogs are caught than mustelids. A few of the rats I remove from the DOC200 stoat traps I throw into the stream for the eels to eat. The native Longfin eel (tuna, ōrea, Anguilla dieffenbachia) is quite common in the local stream
Longfin eel (tuna, ōrea, Anguilla dieffenbachia) get a few rats from council stoat-traps
If you thought that cave wētā (Rhaphidophoridae) were only to be found inside New Zealand caves – Think again. They don’t need caves and they are found all around the world! A new study published in Royal Society Open Science discovered that five geologically young subantarctic islands each have their own amazing and “ancient” endemic cave wētā. [The species themselves are not ancient but each one represents a phylogenetically distinct lineage].
Pachyrhamma longicauda subadult at night on the forest floor.
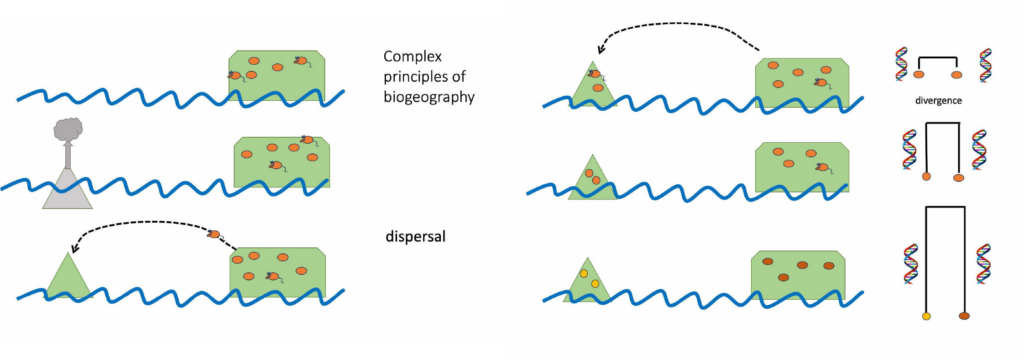
When land emerges from the sea it provides a new home for terrestrial plants and animals. Drifting across the ocean and blowing through the air new organisms arrive. New populations are established and over many generations these populations adapt to their home and might even diversify into a set of new endemic species.
Time series showing organisms colonizing new land and diverging from source population
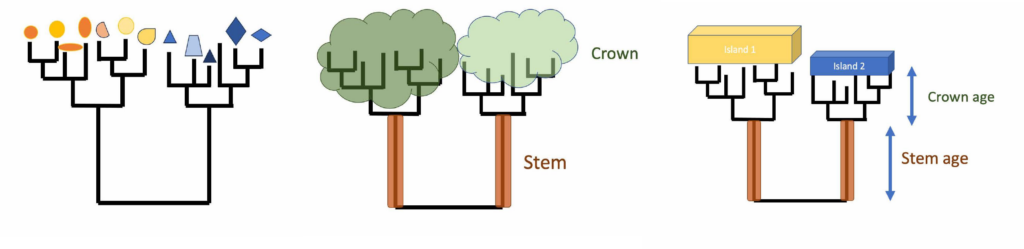
Biologists studying island endemics have long been interested in when it was that islands were first colonised after their formation, and how long it took for novel, distinguishing traits to evolve. A new study of camel crickets on seven subantarctic islands reveals how little we can learn about the time of arrival from the age of the stem. Only the age of the crown group should be used to estimate the speed of evolution.
Phylogenetic tree showing evolutionary relationships, and explanation of the terms stem and crown
The study of Rhaphidophoridae (cave weta/camel crickets) sequenced whole mitochondrial genomes and used fossils from a sister group of orthoptera to calibrate a molecular clock. Species found only on subantarctic islands have sister taxa in New Zealand but the age of the divergence (stem) was found to be much older than the age of their island homes. In all seven cases the island species shared its most recent common ancestor with a sampled relative more than 10 million years ago (most were >25 MYA). This study provides an excellent example of how stem age can mislead biologists into thinking lineages are very old. The reason the stem dates are older than the islands is because close relatives were not sampled. All extant New Zealand genera (with one exception) were included in the study so the lack of close relatives suggests either lack of investment into species discovery or due to extinction. There is still time for us to go looking for close relatives of these island endemics in New Zealand but the islands will be smaller as sea level rises.
the tree on the right shows how extinction has resulted in very distinctive island taxa
Cave crickets are found all over the world including on tiny subantarctic islands. Finding old lineages on young islands shows that these wingless insects are successful at crossing the ocean and colonising new habitat. However, the absence of closely related species elsewhere suggests that extinction is a biologically influential factor with potential to confound traditional biogeographic assumptions
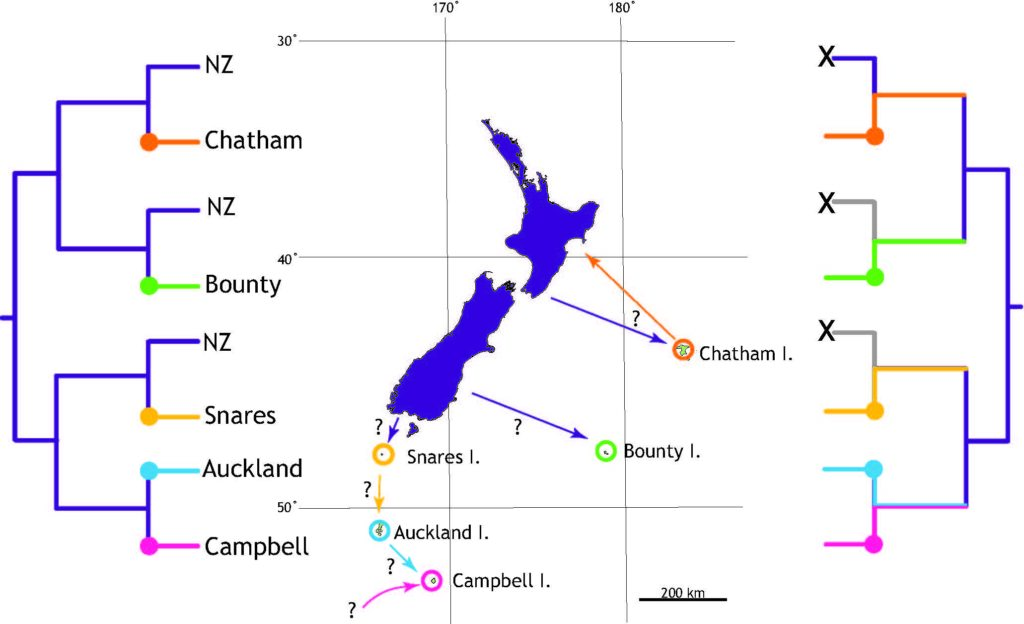
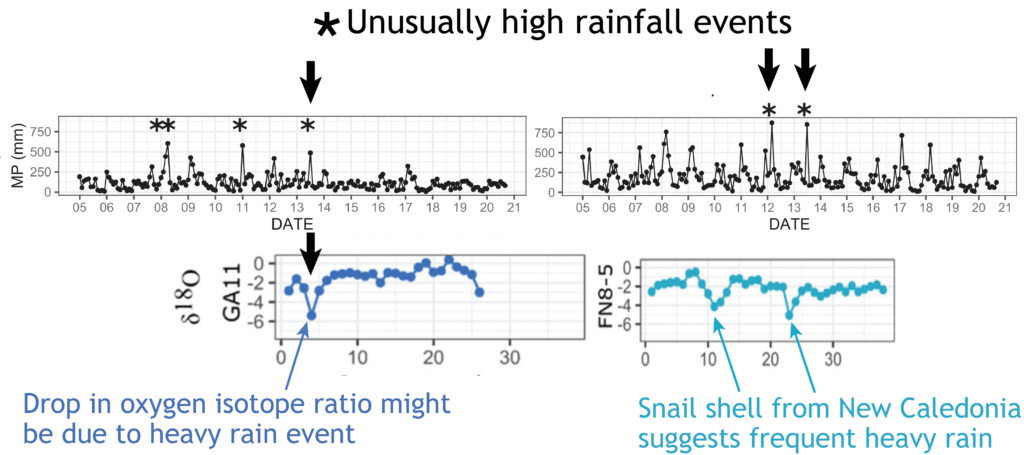
Fossil snails might tell us of the frequency of heavy rainfall in the past
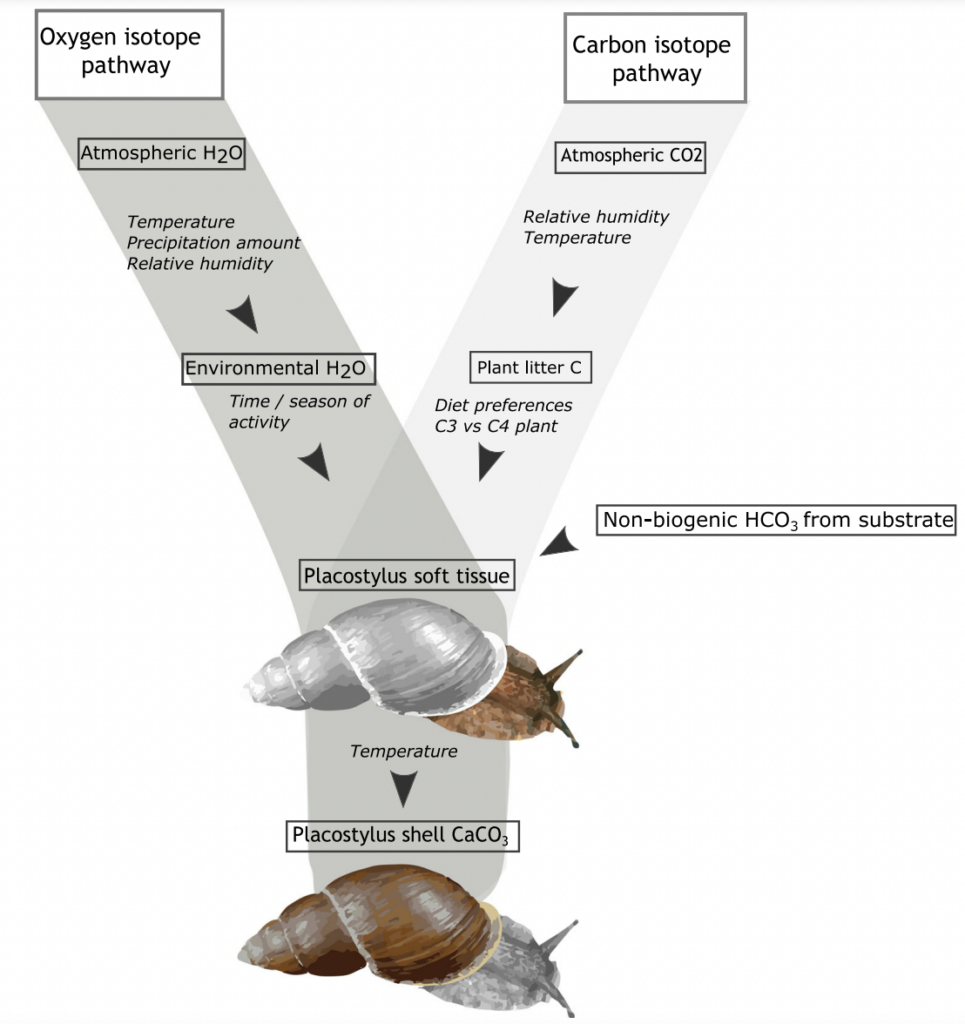
Anew study comparing the stable oxygen and carbon isotope ratios of giant land snails in New Zealand and New Caledonia found a surprising result. New Zealand snails had, on average, higher oxygen isotope ratios values than their counterparts in New Caledonia, counter to the relative isotopic composition of rainwater between these two regions. This research just published in the Journal of Quaternary Science provides baseline data for using the shells of Placostylus snails as environmental proxies – allowing us to use fossil shells to estimate the temperature and rainfall when the snails were alive.
High‐resolution stable isotope profiles from shells of the land snail Placostylus reveal contrasting patterns between snails originating from New Zealand and New Caledonia
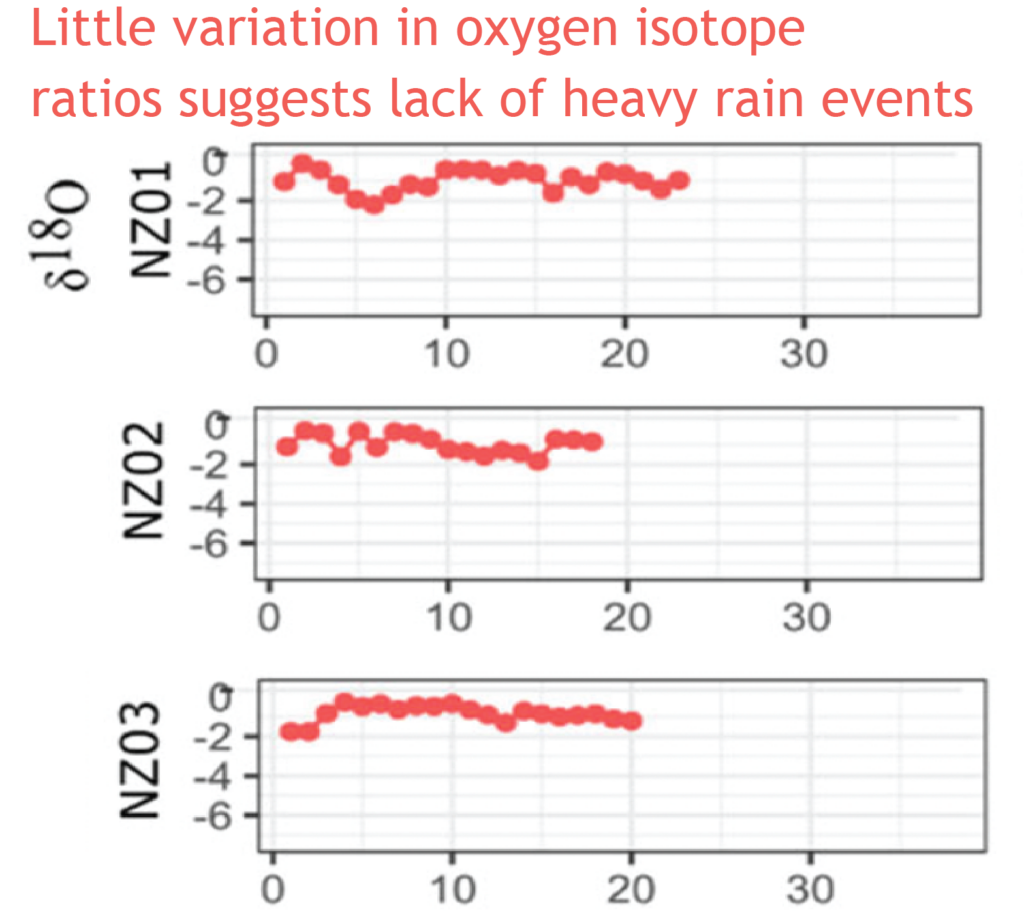
Most interesting are the dramatic drops in oxygen isotope ratios that seem to correspond to heavy rainfall – suggesting an opportunity to dig into the past to compare past precipitation with current frequency of wet weather events in New Zealand. There is also the potential to study the frequency of droughts from the pattern of snail shell growth.
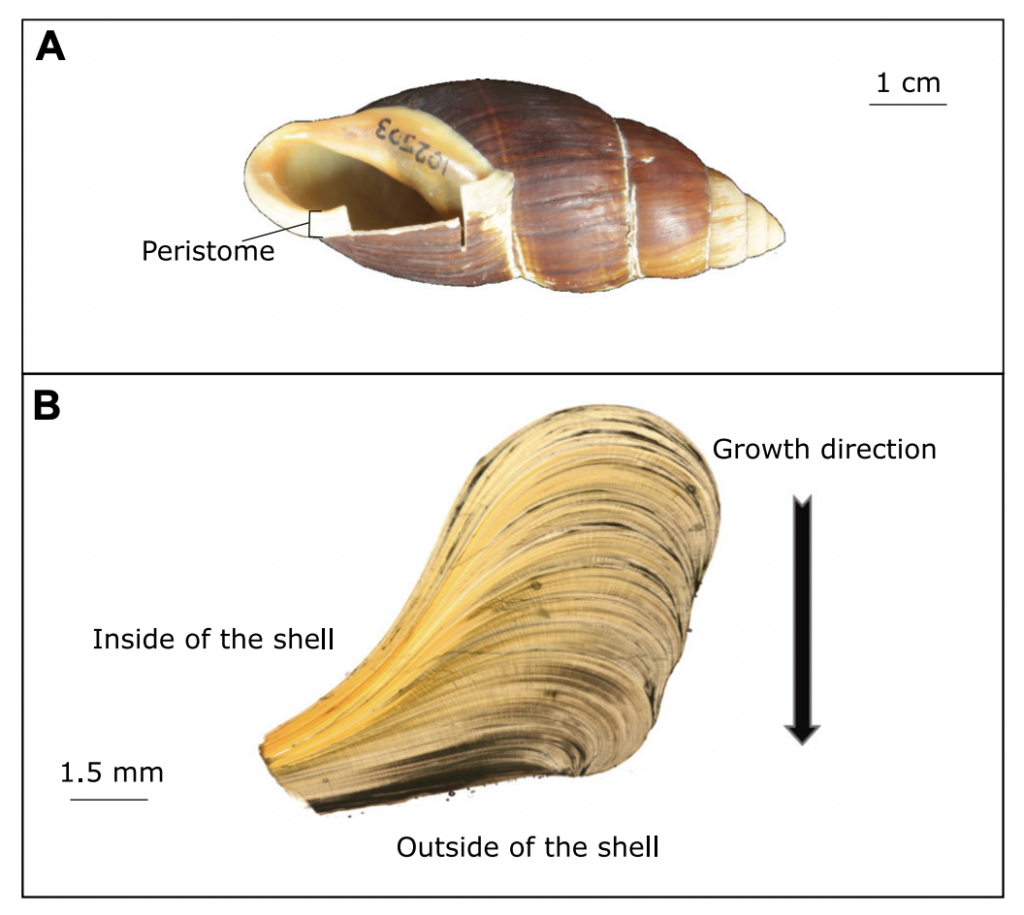
Scientists at Massey University and NIWA sliced up shells of three species of giant land snail (pūpū whakarongotaua; Placostylus). The recent samples from New Caledonia showed drops in isotopic values in their high‐resolution profiles probably linked to periods of intense rainfall.
Very heavy rainfall events produce lower stable oxygen isotope ratios incorporated into the shells of the living-growing land snails. In contrast, the snails from New Zealand varied very little, suggesting that when they were alive, 74 years ago, there were few heavy rain events in the Far North of New Zealand.
The snails (pūpū whakarongotaua; Placostylus) are taonga of Ngāti Kurī who value them as security alarms (the snail that listens for war parties). Ngāti Kurī are working to save the local species from extinction but they are also kaitiaki (guardians) of fossil shells buried in the sand dunes and stored in museums. These fossil deposits could provide information about the past climate through high‐resolution stable oxygen isotope profiles.
“it is exciting to think of all the information locked up in snail shells – the shape of the shell, the DNA and the isotopes can all tell a story about the past” said Mary Morgan-Richards. “As Placostylus snails are slow growing, taking 10 years to reach maturity, and live for a long time, they can each tell their own story. There is much to be learnt by digging into old shells to reveal the frequency of heavy rainfall events in the past.”
What controls the colour of the common mānuka stick insect?
Three stick insects . The same species but different colours living in different environments
When we see how well-camouflaged individual stick insects are, it is hard not to imagine they have chosen either their hiding place or their skin colour, with care. But stick insects cannot see colour – they don’t know if they are green or brown. The reason they are so hard to see is that their cousins who were visible have been eaten. The only ones left are the ones that were not seen by the warblers and silver eyes and chaffinches and tui and sparrows.
Some stick insects can change colour – as demonstrated by the Indian stick insect Carausius morosus (Mangelsdorf 1926), whose colour changes in response to light levels. The trick this insect uses is the movement of pigment granules in it’s skin (cuticle) that alter light absorption and light scattering (Umbers et al. 2014).
Other stick insects like the North American Timema walking sticks have their colour determined by their genes and variation among individuals within the same species is maintained by selection, recombination and mutation (Comeault et al. 2015; Villoutreix et al. 2020). They cannot change colour to suit their environment.
So, what about rō Aotearoa – New Zealand stick insects? No one knows for sure – but maybe you have an idea? I have made some observations of the common mānuka stick insect Clitarchus hookeri and think that genetics is involved.
Colour variation in captivity suggests genetic rather than environmental differences explain this phenotypic diversity
Nymphs of the mānuka stick insect Clitarchus hookeri showing the range of colours a single individual can exhibit at different instars
In boxes at home I allowed green Clitarchus hookeri indivudals from Waikato to mate with brown individuals from Whanganui. I collected the eggs that each female dropped and stored them for a few months until the little stick insects hatched. Nymphs were housed in sibling groups and fed mānuka (Leptospermum scoparium) and climbing rātā (Metrosideros perforate).
All my Clitarchus hookeri offspring hatched as green nymphs. Many remained green as they grew and moulted but some changed colour (see figure). Two males changed to brown when about 30mm long, then speckled grey, before moulting to green as adults, their sisters remained green as they grew. Two females changed to brown when about 25mm long, then pale brown. One matured as a green individual, the other pale brown. Of the twelve stick insects I raised until they were adult eight remained green and four changed colour but only one was still brown when adult. In the next generation, produced by crossing siblings, the similar observations were made; some F2 individuals remained green as they grew, some changed to brown when about 30mm before changing to green as adults.
My observations of the colour of F1 offspring provide preliminary data suggesting that the colour of C. hookeri individuals is genetically determined rather than environmental as nymphs were raised in a common environment. However, I also observed that individuals can be different colours at different stages of their growth. Although changing from green to brown has been recorded in this species (Stringer 1970), changing back to green from brown was unexpected. Individual variation in colour will need to be considered when modelling inheritance and selection of this trait. It is possible that my observations could be explained by relatively few alleles at one or two loci. Further work is needed for us to understand the inheritance of colour traits in the common mānuka stick insect Clitarchus hookeri.
References
Comeault AA, Flaxman SM, Riesch R, Curran E, Soria-Carrasco V, Compert Z, Farkas TE, Muschick M, Parchman TL, Schwander T, Slate J, Nosil P. 2015. Selection on a genetic polymorphism counteracts ecological speciation in a stick insect. Current Biology. 25:1975–1981.
Mangelsdorf AJ. 1926. Color and sex in the Indian walking stick, Dixippus morosus. Psyche. 33:151–155.
Morgan-Richards M, Trewick SA, Stringer IAN. 2010. Geographic parthenogenesis and the common tea-tree stick insect of New Zealand. Molecular Ecology. 19:1227–1238.
Morgan-Richards M, Langton-Myers SS, Trewick SA. 2019. Loss and gain of sexual reproduction in the same stick insect. Molecular Ecology. 28:3929–3941. DOI: 10.1111/mec.15203.
Salmon JT. 1991. The stick insects of New Zealand. Auckland, New Zealand: Reed.
Stringer IAN. 1970. The nymphal and imaginal stages of the bisexual stick insect Clitarchus hookeri (Phasmidae: Phasminae). New Zealand Entomologist. 4:85–95.
Trewick SA, Morgan-Richards M. 2005. New Zealand Wild: Stick Insects. Reed Publishing: Hong Kong.
Umbers KDL, Fabricant SA, Gawryszewski FM, Seago AE, Herberstein ME. 2014. Reversible colour change in Arthropods. Biological Reviews. 89:820–848.
Villoutreix R, de Carvalho CF, Soria-Carrasco V, Lindtke D, De-la-Mora M, Muschick M, Feder JL, Parchman TL, Gompert Z, Nosil P. 2020. Large-scale mutation in the evolution of a gene complex for cryptic coloration. Science 369:460–466.
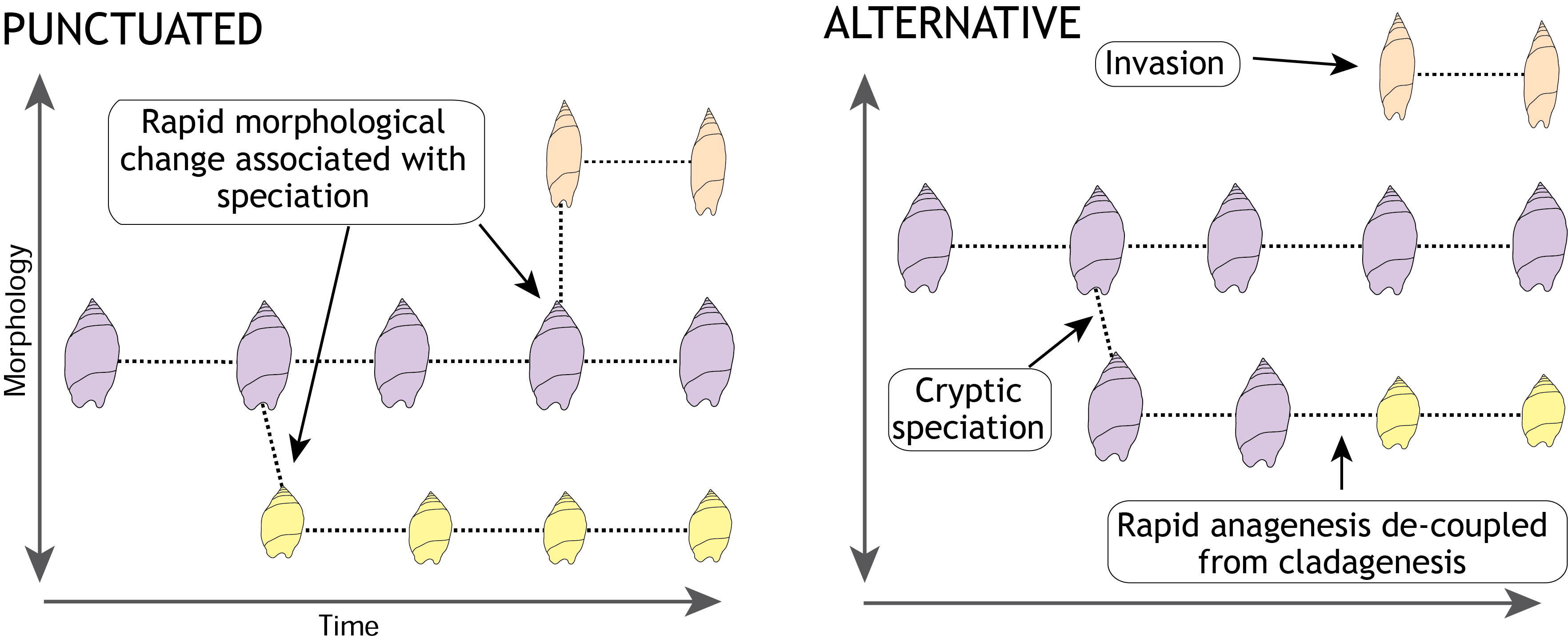
The theory of punctuated equilibrium has two important elements, one is that evolutionary lineages can remain morphologically unchanged for millions of years, the other is that rapid morphological change is associated with speciation.
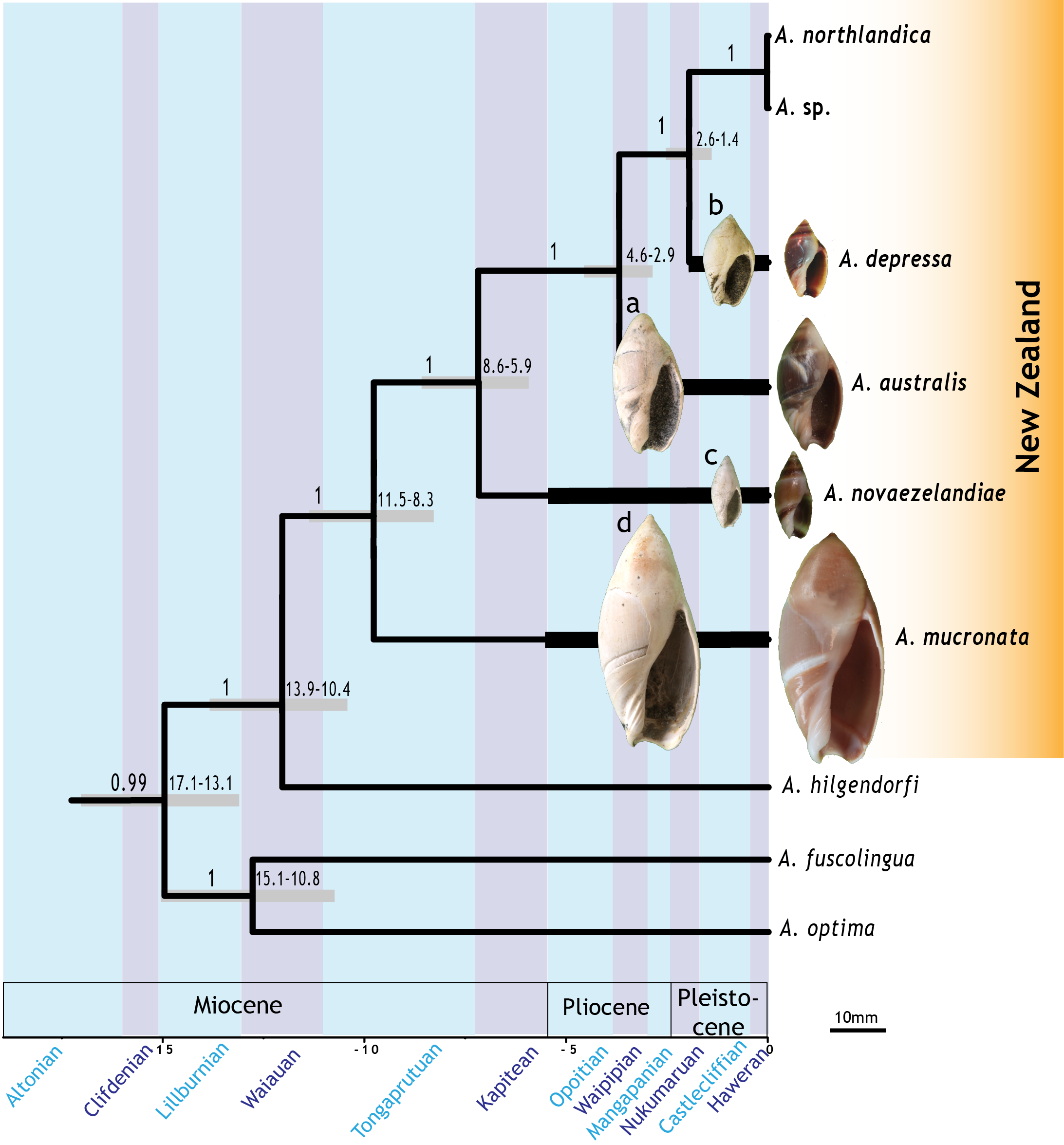
One of the best examples of morphological stasis within biological species comes from the New Zealand Olive Shells (Amalda australis, A. depressa, A. mucronata).
New research on olive shells has just been published: “Phylogenetic topology and timing of New Zealand olive shells are consistent with punctuated equilibrium“.
In this paper we have shown that the three species of New Zealand marine snails (Olive shell Amalda spp) cited by Stephen Jay Gould as important examples of stasis are part of a monophyletic New Zealand clade. This suggests that their evolutionary history has unfolded on the continental shelf around New Zealand, with new species evolving from ancestors in the same region. Using DNA sequences and a molecular clock analysis we determined that lineage splits (speciation) occurred before the 2–3 million years of morphological stasis identified within each of these three species.
Here, we use entire mitogenome and long nuclear rDNA gene cassette data from 11 Amalda species, selected from New Zealand and around the world. Within our sampling, New Zealand Amalda are a natural monophyletic group and estimates of the timing of cladogenesis from the molecular data for the New Zealand group are compatible with the fossil record for extant species and consistent with expectations of punctuated equilibrium.
We have yet to confirm morphological change revealed in the fossil record with the origin of new species, but our study indicates that new taxa in the New Zealand fossil record is not likely to be the result of colonisation by long-distance dispersal of Amalda species from other parts of the world, but instead, represent local evolution.
The punctuated equilibrium model predicts prolonged morphological stasis through time with abrupt change associated with speciation (left). To test this idea we need to show that species diversity is not the result of colonisation from elsewhere or morphological change that is independent of lineage formation (right).
Quote from SJ Gould (1991)
“The best treatment of this objection [that fossil taxa are not biological species] must be sought in studies of living species with good fossil records—where direct surveys can be made for correspondence of a morphological package with a true biological species, and the origin and history of the same package can then be traced in the fossil record and assessed for punctuated equilibrium. I am delighted to report that two such pioneering studies have been published in the past few years, and both support punctuated equilibrium. New Zealand biologist B. Michaux did a morphological and genetic survey of four species in the snail genus Amalda. He found no cryptic populations; each morphologically defined package corresponds perfectly with a biological species. Three of these species extend back in the New Zealand fossil record for several million years. In an elegant, multivariate study of morphological pattern, Michaux demonstrated stasis throughout the ranges of all species. He concludes (in the Biological Journal of the Linnaean Society of London, vol. 38, 1989): This study demonstrates that fossil members of three biologically distinct species fall within the range of variation that is exhibited by extant members of these species. The phenotypic trajectory of each species is shown to oscillate around the modern mean through the time period under consideration. This pattern demonstrates oscillatory change in phenotype [our jargon for overt morphological appearance as contrasted with underlying genetics, or genotype] within prescribed limits, that is, phenotypic stasis.
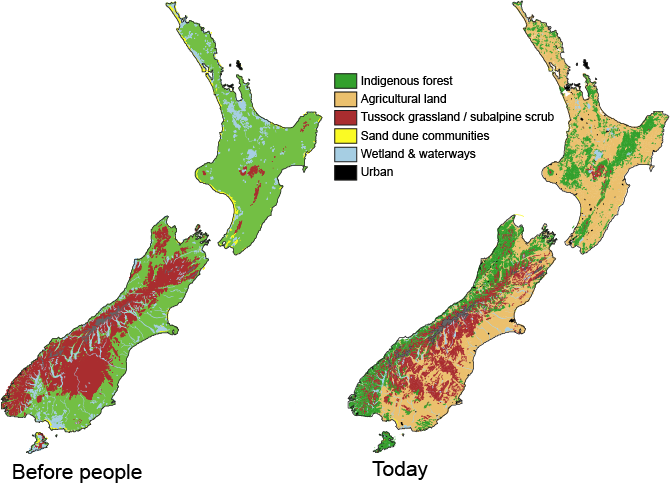
Stand at the side of a mainland island reserve and the impact of humans on New Zealand’s natural environment is obvious. From a landscape naturally dominated by tall forest, agricultural ‘improvement’ rapidly moved us to a uniform, virtual biological desert. Not only are the trees and birds missing, but the lichens, fungi, insects, worms and molluscs are gone. even the bacteria and other microbes of the soil are replaced. In response we resort to counting species and prioritising conservation efforts on the scarcest and restoration effort on the rarest habitats. But, wholesale environmental changes alter no just the abundance of native species but their ecology and interactions. Ultimately, by restructuring the landscape we alter evolutionary outcomes and this has become increasing apparent as research explores biological responses to human induced climate change.
Predator fence on boundary of native forest and exotic paddock. Bushy Park sanctuary near Wanganui.
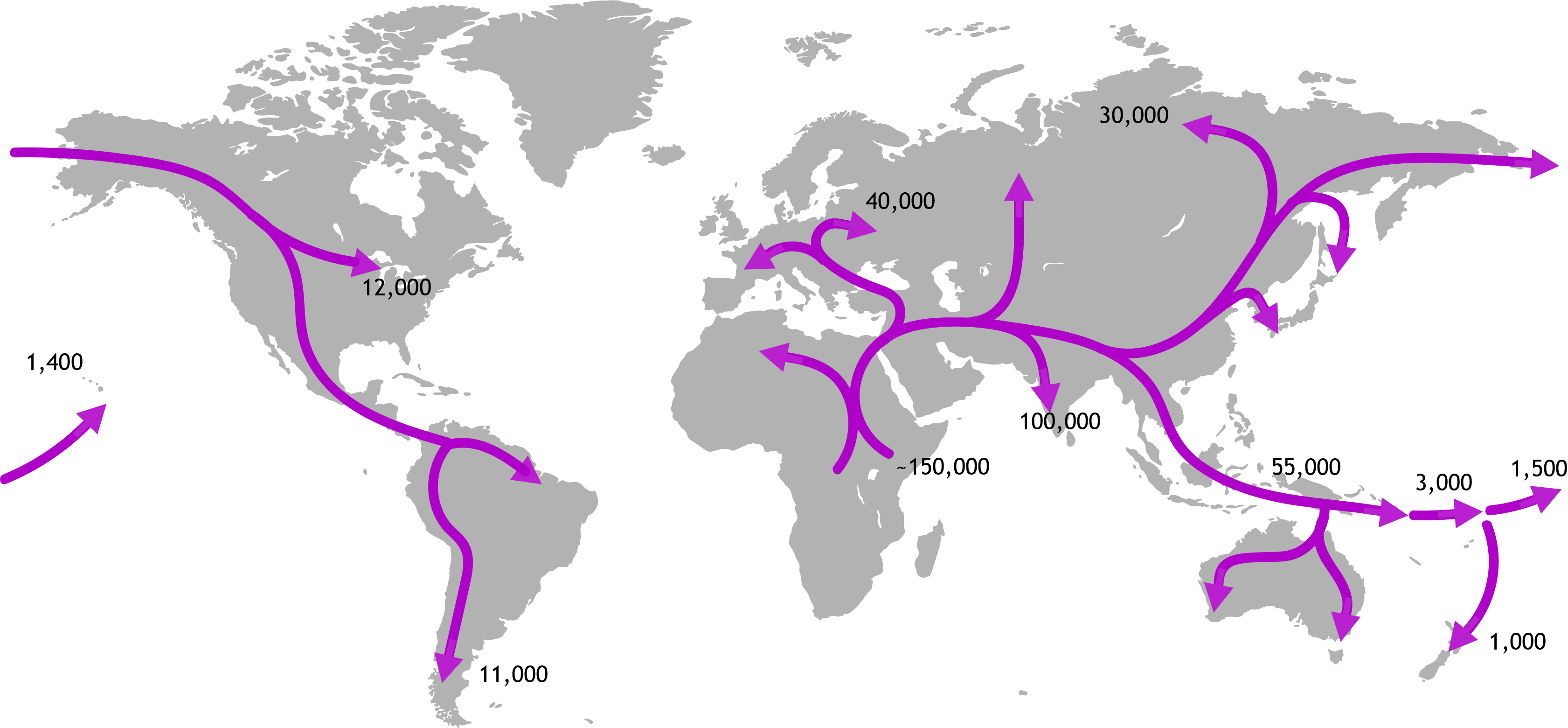
An obvious difficulty with understanding environmental change is that it is much easier to say what is, compared to what was. We are readily inured to the situation and so are accepting of the status quo. One very powerful tool that has helped biologists understand how the geographic ranges of species and population change over time is phylogeography. Simply put, this approach combines information about where individuals and populations of a species are found with information about how those individuals are related to each other. DNA sequence data reveals how closely related individuals are (their genealogy), and how genetically diverse populations are. It is this type of data that shows, for instance, how our human ancestors left Africa and migrated into Europe, then Asia before eventually colonising islands in Oceania. We now know that New Zealand was probably the last major island to have be reached by people travelling by foot and finally boat.
Genetic data from living people has revealed how their ancestors migrated from Africa around the world (values are years before present).
Since the 1990’s phylogeographic studies have revealed the influence of many environmental factors on the distribution of biodiversity. In particular, natural, global climate cycling during the last few million years of Earth’s geophysical prehistory (the Pleistocene epoch) is known to have been influential. We now know for example that in the northern hemisphere repeated extension of the arctic ice cap during ‘glacial’ episodes extinguished populations of all species in northern Europe, Asia and America; remnant populations survived in warmer southern areas. As climate alternately warmed and cooled over 10–100 thousand year cycles, the ranges of animal and plant species expanded and retracted in response.
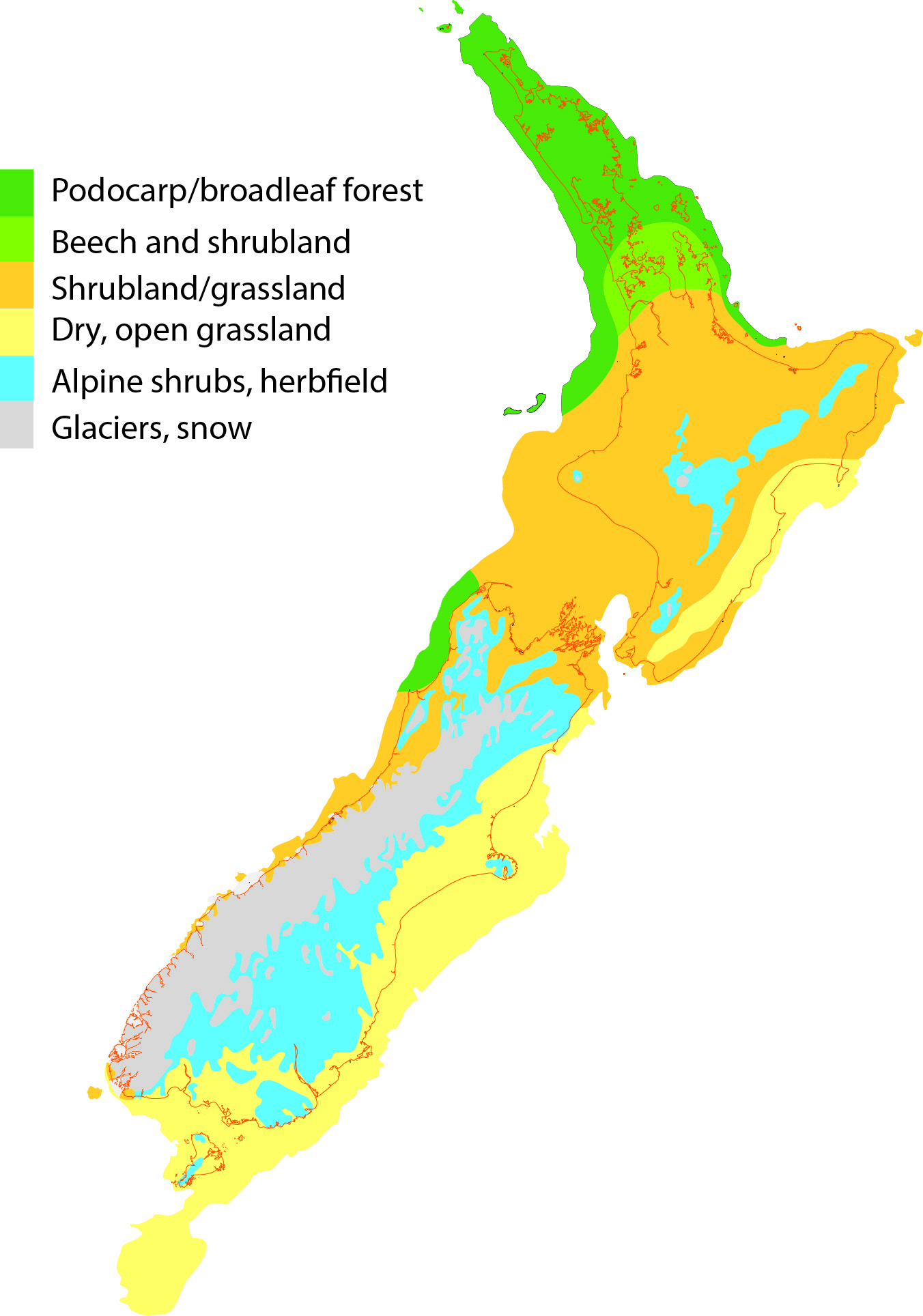
Estimated distribution of vegetation types in New Zealand during the Last Glacial Maximum. See Wild Life New Zealand.
In New Zealand a related pattern of species range change has been inferred. Pollen records show where plant species once lived and genetic data show that during cold phases of the Pleistocene, forest reduced and was replaced in many areas by scrub / grassland communities. Animal species are expected to have responded to these changes tracking their preferred habitat in space and time (or going extinct), and this has been found to be the case for some. North Island tree wētā, for instance, appear to have tracked climate niche.
Phaulacridium marginale
A recent study examined the response of two related grasshopper species. These endemic Phaulacridium grasshoppers live in low elevation habitat, but as is typical of short-horn grasshoppers in temperate regions they require open habitat so they can gain heat by basking in the sun. That means Phaulacridium grasshoppers do not live in forest, and they do not survive above the treeline in the subalpine zone where cool temperatures prevent trees growing (other grasshoppers are adapted to those conditions). So space for Phaulacridium would have been restricted in prehuman New Zealand to scarce open areas such as coastal dunes, river flats, wetlands and semi-arid areas. In fact, one species (Phaulacridium otagoense) occurs today only in the semi-arid McKenzie – Alexandra area of Central Canterbury and Otago. The other species (Phaulacridium marginale) is today found in many places around the country.
Vegetation types across New Zealand before arrival of people (left) and in modern times (right).
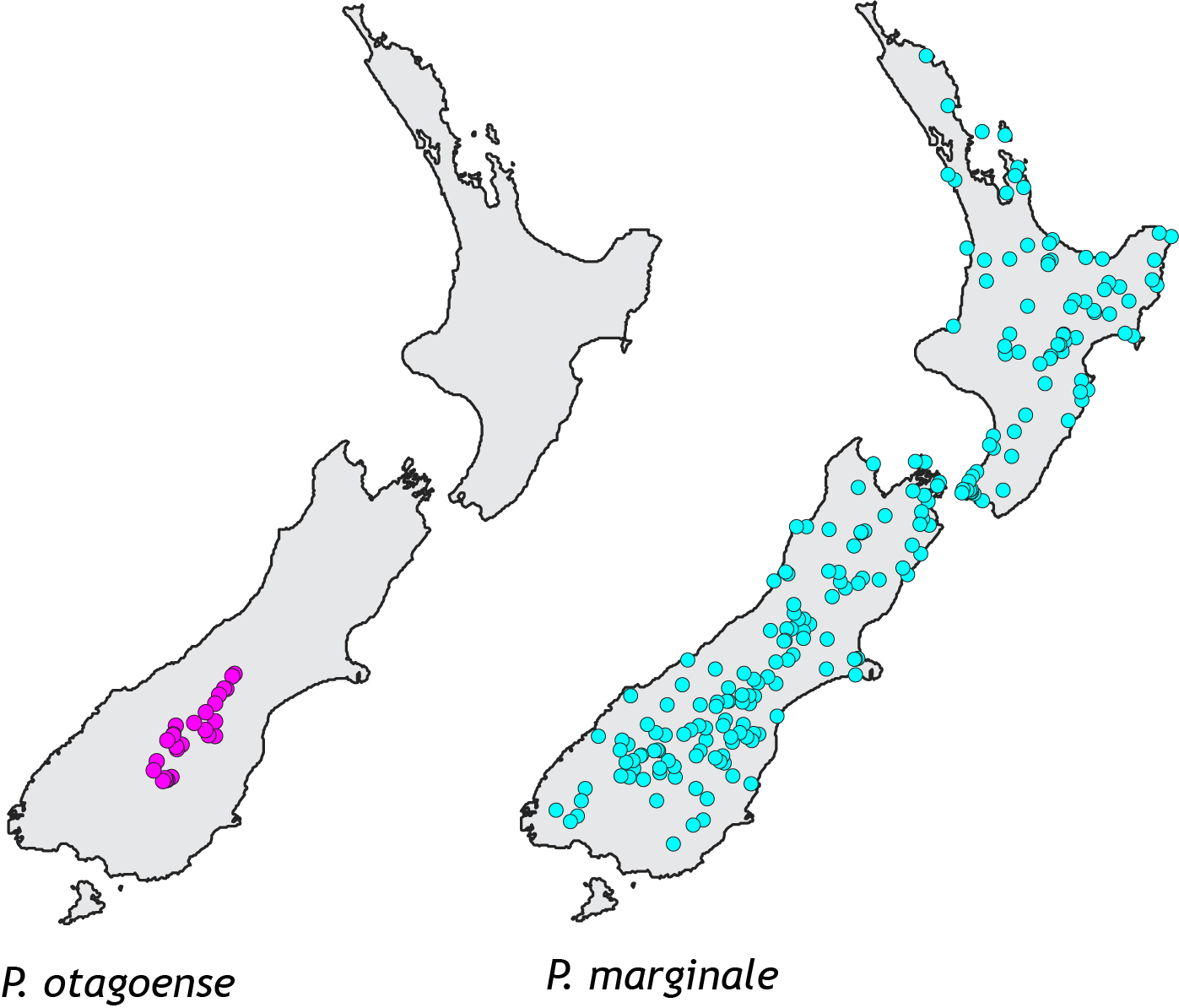
Known occurrences of the two New Zealand Phaulacridium grasshoppers.
A small species range usually means a small population size, compared to a species with a big range; and small populations usually have a lower level of genetic variation. Low genetic diversity is documented in many endangered species such as the famous black robins of the Chatham Islands. Paradoxically, in Phaulacridium the opposite pattern exists; the species with the smallest range (pink in map) has much higher genetic diversity than the widespread more common species. The simplest explanation is that P. otagoense (pink), had until recently a much larger range and so bigger population. Conversely, P. marginale (turquoise) appears to have expanded its range recently and has not yet had time to accumulate new genetic diversity.
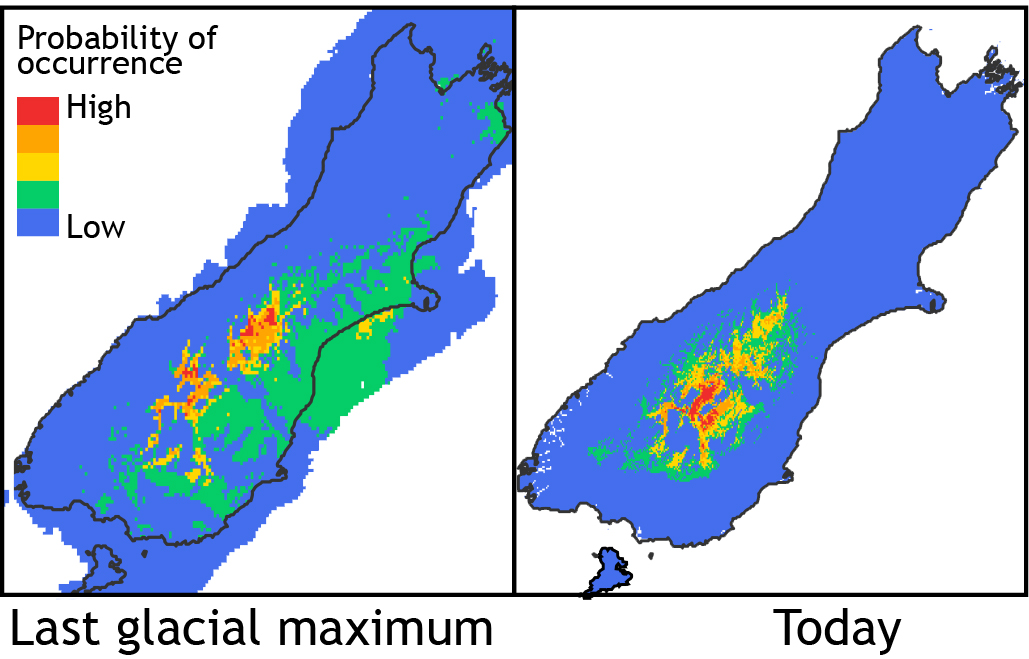
Niche models for Phaulacridium otagoense indicating optimal habitat (red, orange) during the last glacial phase may have been similar to today.
It is known that global temperatures had recovered from the last cold phase of the Pleistocene by about 15,000 years ago. Perhaps P. otagoense had a much larger range in the period before that when cooler, drier conditions allowed scrub grassland to expand; similar to conditions where it occurs today? Niche modelling indicates that in current conditions the potential range of this species is bigger than the actual range in which it is found, and taking into account estimated temperatures during the last glaciation suggests that the habitat preferred by this species had not been much more extensive.
So, probably the major change in fortunes for these Phaulacridium species relates mostly to the recent expansion of P. marginale. Climate modelling shows that the range of this species today is close to the potential occupiable range, but there is a problem. Although the climate across much of New Zealand suits this grasshopper, other factors in the environment do not. In particular, the presence of native forest excludes these little grasshoppers because they need to bask in the sun every day to warm up. How has P. marginale become so abundant and widespread?
The answer lies not in global climate change, but in recent anthropogenic changes to the environment much closer to home. By removing New Zealand native forest, humans created a landscape with the climatic conditions to allow P. marginale to increase in abundance and expand its range across the country. The addition of a mix of northern hemisphere grasses and herbs that thrive in this artificially open environment provided the nutrient-rich food for P. marginale. So that’s great! Well no.
Males and females of different species are capable of reproduction when they meet due to anthropogenic habitat change, resulting in loss of diversity. Male Phaulacridium otagoense with female P. marginale.
The increase in available habitat has meant that the spatial range of P. marginale now meets the range of P. otagoense. Where they meet, the grasshoppers makes mistakes when choosing mates resulting in gene flow. Genetic evidence shows that pure P. otagoense remain in only part of their natural ecological range. Genetic mixing is part of the natural evolutionary mill, but around the world human activity has accelerated the rate at which species meet and interact in new ways. This adds to the trends of biological homogenisation that characterises biodiversity loss in the Anthropocene.
New Zealand stick insects have invaded the United Kingdom, but in the process they have lost the ability to reproduce sexually. This is odd because the vast majority (more than 99%) of multicellular creatures (primarily eukaryotes) engage in sex during reproduction.
Sex involves two individuals with different properties. Typically one sex (the male) produces abundant small and often motile gametes that carry genetic information to the larger egg produced by the other (female). Through this process, genetic information is passed from two parents to their offspring and results in shuffling of genetic variation. The results are readily evident in the variation seen among offspring that is prominent in human families.
Stick insects are (mostly) no exception even though scientist can show that reproduction without two sexes can have a numerical advantage over sexual reproduction. Simply, females that make only self-fertile daughters leave more of their genetics to future generations. Theoretically it seems that clonal reproduction is advantageous, as long as the environment does not vary too much; producing offspring that are not the same as the parent could make some of them less successful. It is telling then, that despite the numerical advantage of clonal reproduction, that vast majority of large organisms do use sexual reproduction. Natural selection has made its choice.
One group of New Zealand stick insects includes individuals that differ in colour, size, and shape. In particular the number and size of spines they have varies among individuals. This group (genus Acanthoxyla) includes several described species, although in this case defining species is difficult. All are female, which means all come from self-fertile eggs produced by one parent (the mother). Hatchlings grow up to look like their mums, so are effectively clones.
Among the many individuals of common and widespread Acanthoxyla (literally: prickly stick) observed in New Zealand, no male has been encountered. Yet. But recently a male belonging to this genus turned up in England.
Rare males like this emerge among all-female stick insect populations, probably as a result of a random mutation deleting one of the XX sex chromosomes that denotes a female stick insect. XO individuals are male in appearance, but are usually not reproductive.
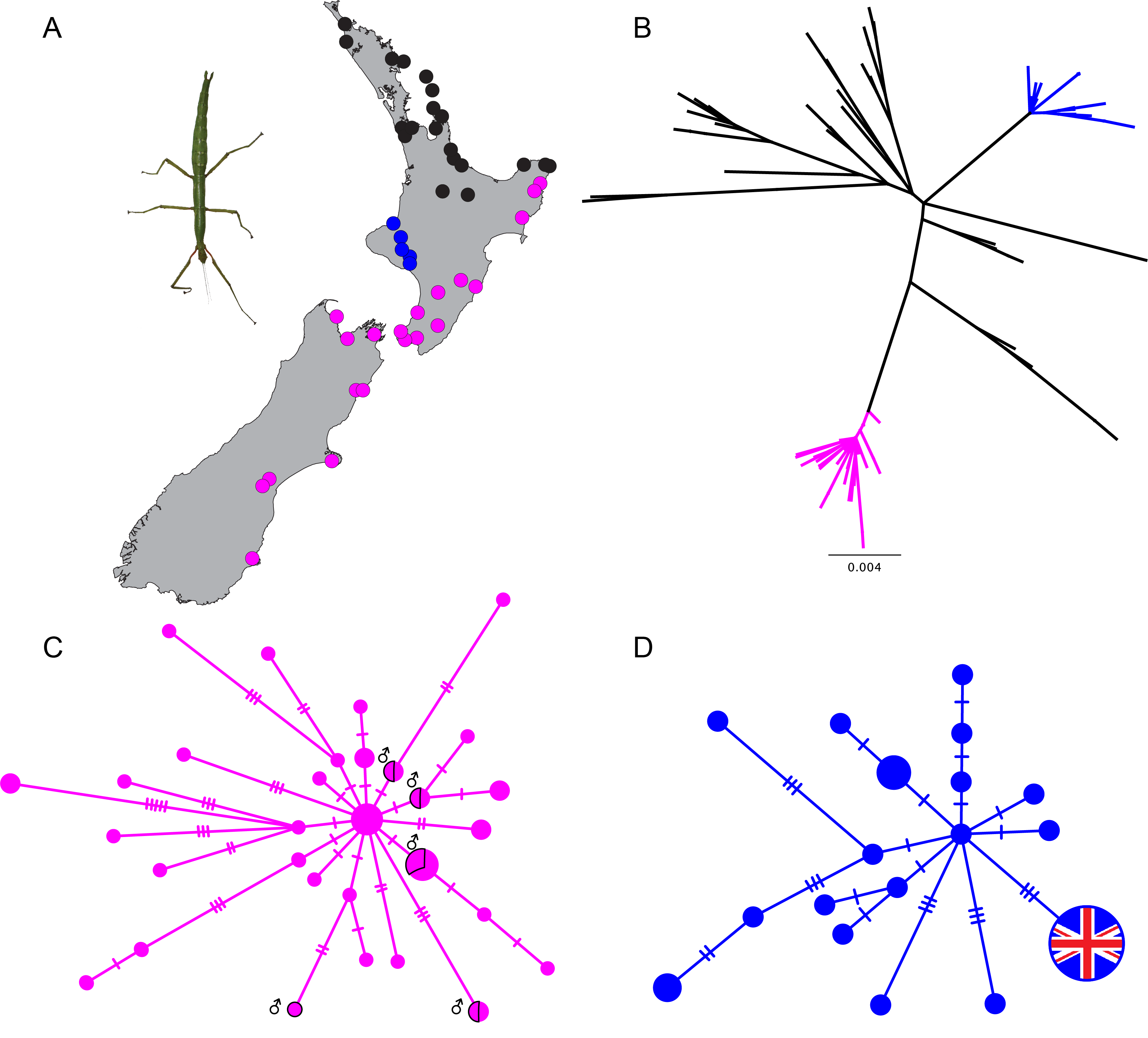
Research on the New Zealand genus Clitarchus has been revealing about the switching between sexual and asexual (all female) reproduction. As reported in Nature a population of Clitarchus hookeri accidentally introduced to the UK about 100 years ago has lost not only its homeland but also its sex life.
Analysis of genetic variation shows that the origin in New Zealand of the UK stick immigrants was most likely in Taranaki, North Island. This agrees with historical records indicating that native plants collected in this area were shipped to England and then the nearby Isles of Scilly. In particular the Abbey Gardens on Tresco are now home to a range of New Zealand plants, and it is likely that stick insect eggs in the soil around plant specimens were accidentally transported around the world. Hatchlings that grew into adult stick insects able to produce abundant self-fertile females were likely at an advantage. The potential of this species to switch to asexual reproduction has also resulted in a pattern of geographic parthenogenesis in New Zealand.
Genetic variation (mtDNA COI) in Clitarchus hookeri across New Zealand (A, B), highlighting the mainly parthenogenetic lineage in NZ (C), and the lineage associated with the one variant found in the UK population (D).
Closer examination of two New Zealand populations of the same species add to our understanding of the drivers and mechanisms of reproduction strategy switching. The UK population lost sexual reproduction and evolved a barrier to fertilisation, which has been demonstrated by providing captive female stick insects from UK with NZ males. Meanwhile two NZ populations recently gained sexuality and genotypic data indicate this happened via two different pathways.





















 One group of New Zealand stick insects includes individuals that differ in colour, size, and shape. In particular the number and size of spines they have varies among individuals. This group (genus Acanthoxyla) includes several described species, although in this case defining species is difficult. All are female, which means all come from self-fertile eggs produced by one parent (the mother). Hatchlings grow up to look like their mums, so are effectively clones.
One group of New Zealand stick insects includes individuals that differ in colour, size, and shape. In particular the number and size of spines they have varies among individuals. This group (genus Acanthoxyla) includes several described species, although in this case defining species is difficult. All are female, which means all come from self-fertile eggs produced by one parent (the mother). Hatchlings grow up to look like their mums, so are effectively clones. Among the many individuals of common and widespread Acanthoxyla (literally: prickly stick) observed in New Zealand, no male has been encountered. Yet. But recently a male belonging to this genus turned up in
Among the many individuals of common and widespread Acanthoxyla (literally: prickly stick) observed in New Zealand, no male has been encountered. Yet. But recently a male belonging to this genus turned up in