
In this section we consider :
i) C (and N) sequestration following ‘intensification’ (meat to dairy; fertiliser, irrigation)
ii) C (and N) sequestration following shifts between high and low N inputs, using dairy or meat
i) C (and N) sequestration following a shift from low input meat to higher input dairy
The effects of numerous changes in management on the amount of carbon (C) sequestered in the system (predominantly in soil organic matter) are evident in all the associated pages ‘supplements’; ‘irrigation’ (eg) where outputs are presented as response graphs (eg see Figs (b) under those pages).
But concern has arisen over how changes in land use, over the last 20-30 years in NZ, and which involve multiple changes in inputs, will have affected the amount of C sequestered.
Here we describe the time course of changes in the amount of C sequestered, and consider explanations for these, following a ‘characteristic’ shift from a low input ‘meat’ system, to a ‘dairy’ system, receiving the same, or higher, N inputs; and with (and without) summer irrigation.
Background: observations and explanation from a major field survey of soil C
In a recent paper, Mudge et al. (2016) presented empirical data from long term field observations in which it was possible to compare ‘paired’ fields/farms in which one ‘system’ had been irrigated, the other not (and for areas under dairy in particular) … and the data is used to conclude that there may in general be a decline in the amount of C (and N) sequestered in soil organic matter, as a consequence of irrigation. The work is seminal, and the paper is exemplary, in what it describes. The data focus on analyses of the soil under the contrasting ‘treatments’. The paper relays that there may be some surprise that irrigation (that should stimulate plant growth and so C fixation) could lead to a reduction (lessening) in the amount of C sequestered, and in its Discussion the paper offers a hypothesis… that the decline in soil C was due to water stimulating (microbial) soil respiration.
Below, we consider responses to water, nutrient (N) inputs and animal class, using the HPM, to test whether it is necessary to evoke such a non-linearity, and/or an undescribed response by the soil microbial population, to account for the phenomena observed in that study. That is not to say that the hypothesised response is challenged, noting respiration was not measured in that paper/trials, but simply whether there are other alternative explanations that require us to incur no processes beyond what are already well established.
An alternative explanation:
Our alternative explanation is that the decline in soil C (indeed all C sequestration) is driven by the way adding water can lead to nitrogen limitation. This in turn reduces C inputs and so the capacity to sustain the amount of C sequestered.
We argue that the major impact of irrigation is first and foremost to remove physiological constraints to C fixation (photosynthesis), and so increase plant growth (and N uptake) during periods of otherwise water deficiency. In this sense, plant growth responses to water are similar to responses to N. Both stimulate plant growth, though the extent of the response to either would depend on which (or both) of water, or N, are currently limiting. Hence these two ‘limiting factors’ themselves interact.
Note, in addition, water input can stimulate the seasonal total of plant growth in that it, effectively, ‘adds time’ .. that is, it allows more photosynthesis, and so growth, to take place during months of ‘water-deficiency’ in summer, and so adds C capture and plant growth, notably at a time of high light energy receipt. In the absence of irrigation, C capture and growth during that period would have been restricted.
But this creates a dynamic interaction between water and nitrogen, as a consequence.
Any increase in plant growth that does arise, following irrigation, should sensibly lead to an increased removal of herbage (hence both C and N), from the system, in animal products.
Hence the addition of water (irrigation), by increasing the amount of N removed in products, has the potential to lead dynamically to growing nitrogen deficiency (creating a nitrogen limitation). That nitrogen deficiency would limit C fixation, and so in turn limit the capacity to sustain the amount of carbon sequestered. The likelihood of a building N deficiency will depend critically, of course, on whether the use of irrigation were accompanied by a sustained increase in eg fertiliser N input rates.
In short, we propose that water input can lead to a nitrogen deficiency, and in that way decrease the amount of C that is sequestered.
This hypothesis is summarised below:

To examine the alternative explanation…
Time course
First we plot out the pattern of changes over time in the amount of C sequestered in the system (predominantly, but not exclusively, in soil organic matter) as predicted by our dynamic modelling. This graph depicts changes in the amount of C (a state, not a rate) following a shift from a low input ‘meat’ system (M60) to various ‘dairy’ (L..) systems options (as explained below).
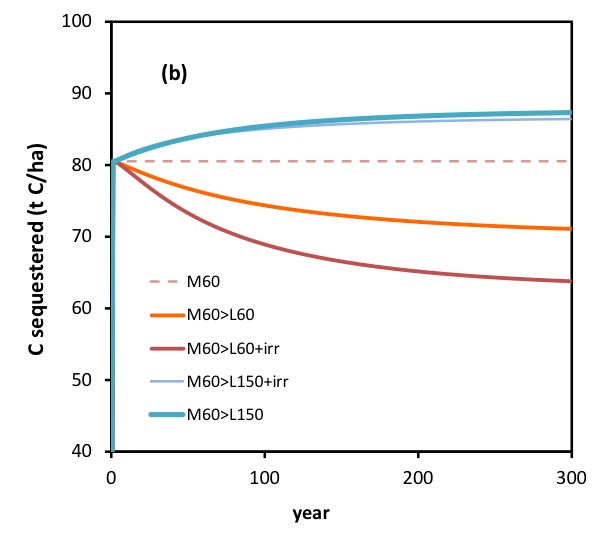
The time course reveals that the amount of carbon sequestered can decline substantially following a shift from a low N input ‘meat’ system (M60) to a ‘dairy’ system at that same low N input rate (M60>L60), and that the decline is even greater if accompanied by the use of summer irrigation (M60>L60+irr). However, no decline is observed, indeed the amount of C sequestered increases, if the shift from low input ‘meat’ to ‘dairy’ is accompanied by an increased input of fertiliser N (M60>L150), and there is no significant worsening of the accumulation of carbon if that shift also involves irrigation (M60>L150+irr). These patterns of changes are not unlike those observed by Mudge et al (2016).
These pattens of responses are also consistent with how it may be a growing deficiency for N that is limiting C sequestration, in that adding a higher rate of N inputs reduces the prospect for a loss of sequestered carbon. The pattern of responses, in graph (b), follows N inputs and not that of irrigation per se.
But much further explanation/evidence is required:
The changes in the amount of carbon sequestered, are shown below, in the context of changes in yield of products (a); total N releases to the environment (c); and methane emissions (d) (all per ha). (Note: graph (b) is a repeat of that shown larger above)
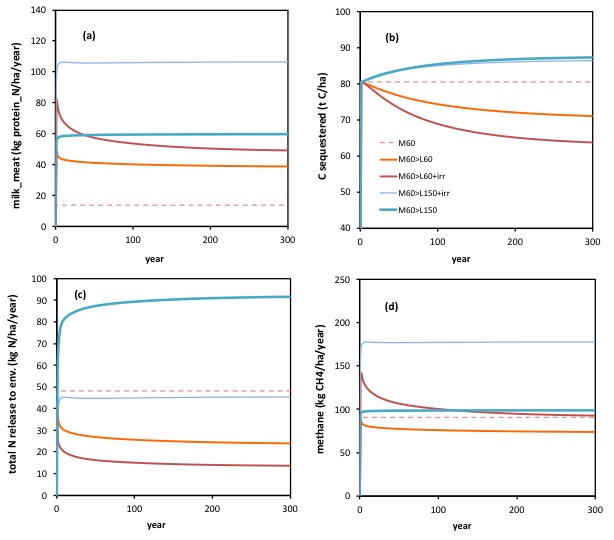
In (a) we can see that in all cases the shift from low input ‘meat’ (M60) to ‘dairy’ (all ‘L’) involved a major increase in production, and hence, note, in the removal of N from the system in products. The units of production here, graph (a), are indeed in kg N/ha/year. The increase in the removal of N in products (yellow line graph (a)) is reflected in a decrease in the release of N to the environment (yellow line graph (c)). What is sustainably removed in products cannot be there to be released to the environment.
Where irrigation was applied, but where N inputs were not increased following the shift to ‘dairy’ (M60>L60+irr; see red line), note how production (and the removal of N in products) was initially increased, but that initially elevated rate of removal of N could not be sustained. This too is in keeping with the notion that irrigation alone will lead to increased plant growth, but so also to a greater removal of C and N in products. Note too how the total release of N to the environment, see (c) was lower as a consequence of the greater removal of N in products (at a given sustained N input rate).
Where the shift to ‘dairy’ was accompanied by an increased input of N (up from 60, to 150 kg N/ha/year), hence from ‘M60>L150’ there was a sustainable increase in the removal of C and N in products (see thicker blue line, in (a) ), but this was accompanied by a major increase also in the rate of release of N to the environment (see (c)).
Where the shift to dairy was accompanied also by the addition of water (irrigation in dry summer months) hence ‘M60>L150+irr’, there was an even greater stimulus to the amount of (C and) N removed in animal products (see thin blue line in (a) ), but this time a much lower release of N to the environment (see (c)).
Hence very different combinations of yield in animal products, and N releases to the environment, can be achieved for the same outcome regarding C sequestration (in (b)) , depending on whether adding fertiliser N is associated with also adding irrigation. Again, a dynamic interaction, as proposed.
In short:
The amount of carbon sequestered, as shown above, only decreases where the use of irrigation had not been accompanied by an increase in N fertiliser inputs, to compensate for the greater removal of N in milk that the irrigation had stimulated.
Is soil carbon loss due to water inducing increased soil respiration?
This seems unlikely, in that (as shown below) irrigation, during the critical summer months, led first and foremost to increases in photosynthesis and so greater inputs of carbon to the system. These greater rates of input of carbon mean the system can ‘support’ , indeed makes inevitable, the higher rates of soil (and whole system) respiration.
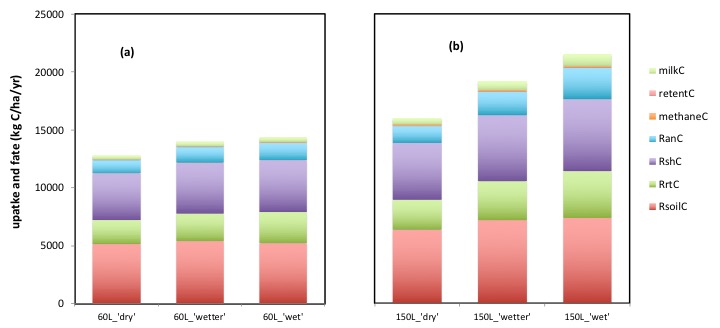
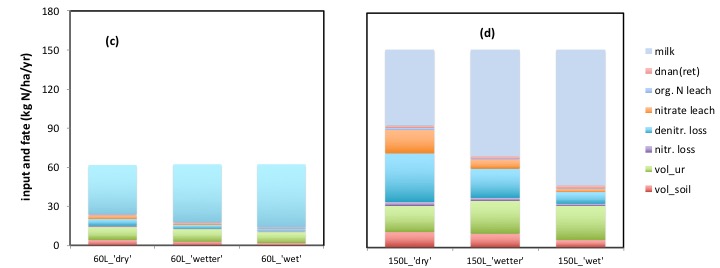
The graphs (a) and (b) show the effects of 3 levels of irrigation (‘dry’ = none; ‘wetter’ = 50% increase in water, and ‘wet’ = a doubling of water input, in critical summer months) in a ‘dairy’ system (‘L’) either at a sustained 60 kg N/ha/yr, or at a sustained 150 kg N/ha/yr.
The top of each bar is the total input of carbon to the system in (‘gross’) photosynthesis, and within each bar is shown the breakdown, or fate, of that input of carbon, between the multiple ‘outputs’ of carbon from the system.
The breakdown of the fate of carbon reveals, strikingly, the scale of the ‘loss’ (release) of carbon in respiration. The most substantial release of carbon is from soil respiration (includes litter) and that from roots (bottom two categories). Adding in the respiration from the plant shoot (third category up from bottom), allows us to see the total respiratory loss of carbon from the plant. Above this is the loss of C in respiration from the animal, and the tiny (quantitatively in this context), loss of carbon in methane, retention and milk.
Returning to our ‘alternative hypothesis’, irrigation (and fertiliser N inputs) both increased the total uptake of carbon in photosynthesis (the top of each bar). The increase in carbon capture due to water input, was limited, at 60 kg N/ha/yr fertiliser N input; the increase in carbon capture due to water input was greater, at 150 kg N/ha/yr… hence the water and N responses interact, as proposed.
Irrigation, and fertiliser N inputs certainly did lead to an increase in ‘soil’ (indeed all) respiration, but the increases in respiration are, very evidently and inevitably at a sustainable steady state, totally supported by the corresponding increases in total C inputs.
Moreover, (see below), it is clear that these increases in respiration, despite taking place, are not inevitably associated with, nor cause, a decrease in the amount of carbon sequestered. In graphs (e) and (f) the amount of carbon sequestered was greater (in (f)) where respiration (see (b above) was greatest.
So, respiration was in all cases increased by irrigation, but almost exclusively in that irrigation had increased the total capture (input) of C, and so there was more carbon to be respired as a consequence. There was also greater biomass (eg in soil) to give rise to that greater respiration.
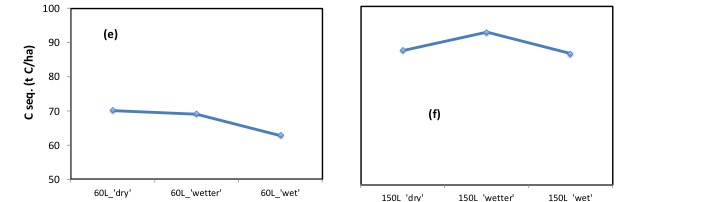
This strongly suggests that the losses of sequestered carbon seen in empirical studies, following irrigation, are not the consequence of an increase in soil respiration. (respiration did increase, but so did soil C sequestration).
N balance reveals how irrigation increases the removal of N in products, and so reduces N releases (loss)
If we turn to the N balance (input and fate of N) in the same examples, there is a very distinct contrast between how N fertiliser input, and water input, interact to affect the N cycle, compared to how they affect the C cycle.
The top of the bars, in (c) and (d) below, reflect the total N inputs to the system, and so reflect the two different N input rates (60 kg N/ha/yr and 150 kg N/ha/year). hence the total amount of N to be accounted for increased substantially, and simply, with N inputs. The effects of adding water (summer irrigation) is entirely on the relative fate of that N, and so the balance between the multiple processes of N release to the environment (and atmosphere).
In graph (c), and so at 60 kg N/ha/day, there was only a limited increase in the rate of removal of N in milk (as explained above). This had relatively small effects then on the fate of N, both in absolute terms, and in relative terms. The bars in graph (c) below show the breakdown of the release of N to the environment by multiple routes, as well as how the addition of water, reduced overall N releases, by increasing the amount of N removed in animal products.
In graph (d), and so at a higher 150 kg N/ha/yr N input, the response to water in terms of the removal of N in animal products, was greater (plant growth less N limited). Due to the substantial increase in C capture following the addition of water (as seen above) a far greater rate of removal of (C and) N, as milk, is incurred and could be sustained.
Note how, given the fixed rate of N inputs, and yet a far greater removal of N in products, under irrigation, the absolute rates of most major N releases, eg denitrification, and potentially also leaching, may actually become smaller under irrigation, per se, than in un-irrigated areas. For certain, higher water content in soils (low oxygen content) would itself favour denitrification (the process as well as the release of N associated with it). That is not denied here. What remains however is to note that any process that depends on the amount (and sustained rate of replacement) of N available as a substrate, cannot be sustained at a high rate, regardless of how suitable surrounding conditions (low oxygen) are, when the amount of substrate (and replacement rate) is sustainably reduced in absolute terms. Hence despite the soil being ‘wet’, denitrification and even leaching, can be reduced under irrigation.
The situation where BOTH irrigation and nitrogen inputs are increased is NOT overlooked here. This can be seen by eg comparing 60L_’dry’ with 150L_’wet’.
Carbon losses closely follow (track) pattern of C inputs
A further indication of how it is the changes in the rate of input of C (C capture in photosynthesis) that determines the amount of C that can be sustainably sequestered, (and not a factor affecting the rate of loss of C in respiration) is to compare the changes over time in the rate of inputs of carbon (below left) and the rate of outputs of carbon (below right). The rate of outputs of carbon, recall, are very predominantly ‘respiration’.
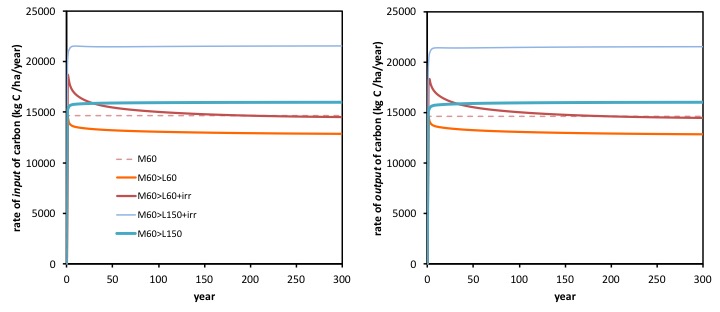
These two graphs reveal how irrigation and fertiliser N treatments did lead to changes in the rate of carbon outputs (right hand graph), but these closely follow, year by year, changes in the rate of inputs of carbon (carbon capture in photosynthesis) as seen in the left hand graph.
The time course of the rate of sequestration of carbon within the system (and predominantly in soils) is the difference, year by year, between the rates of inputs and outputs in the graphs above. We can plot this as below (top graph) along with the equivalent rate of sequestration of nitrogen (bottom graph).
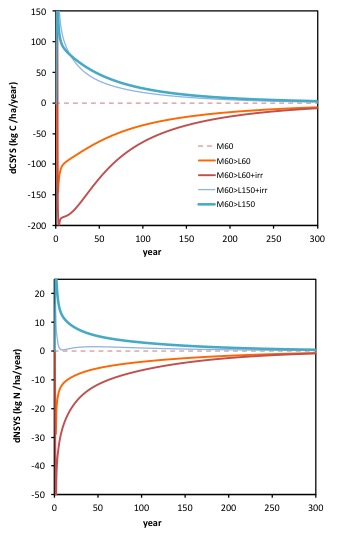
The top graph shows the pattern of changes in the rate of sequestration (hence dCsys/dt) , that led to the changes (increase or decrease) in the amount of carbon sequestered (C).
Below this we see the corresponding changes in the rate of sequestration of N (dNsys/dt) in the system. Note both C and N sequestration approach zero as the new dynamic steady state is approached, albeit taking some 50 to 100+ years.
Most important is to note how the changes in the rate of nitrogen sequestration in the system (lower graph) precede the changes in the rate of carbon sequestration in the system (top graph). In dynamic systems, this would be regarded as evidence that it was the changes in the N cycle that were driving the changes in the C cycle, and so again, that these phenomena do not support the hypothesis that the reduction in the amount of carbon sequestered following irrigation, was a consequence of irrigation increasing soil respiration. It supports, rather, the alternative explanation, that the loss of sequestered carbon that can be seen following irrigation, is the result of a growing nitrogen deficiency, induced by the increased removal of N in products, that irrigation made possible.
In conclusion: …
There is no denying the substantial contribution to recognising the effects of irrigation and land use change on soil C nd N sequestration, made by the paper of Mudge et al (2016). That paper superbly collates and describes empirical observations, ‘real data’, of how C and N sequestration could be reduced in some circumstances, while in others less so, following irrigation. Their paper discusses a possible explanation, that increase in soil water prompted increased soil respiration, and this led to a decline in soil C (and with it soil N). This is totally plausible. The phenomena (increased soil respiration) were not however measured, and the explanation requires evoking a mechanism, and non-linearity, that has not been demonstrated as being causal.
Our alternative explanation produces phenomena that match closely those changes in soil C and N, observed by Mudge et al (2016) including an associated (but for us a resultant) increase in soil respiration following irrigation and land use change. But it is simpler in that it explains changes in the amount of C sustainably sequestered, in terms of the changes in the amount of carbon captured (hence the rate of inputs of C) by the system. Ultimately losses of carbon (respiration) cannot sustainably exceed the rates of inputs of carbon (photosynthesis) and so we are confident of that generality. This does not deny that there may well be complex non-linearities, in how water inputs, interacting with fertiliser N use, may prompt multiple changes in the soil organism community, whether archeae, fungi, bacteria, or higher organisms, and these alone may alter the balance of soil C and N processes. For now, we propose only that there are alternative, arguably simpler, explanations for what was observed by Mudge et al.
The comparison offers a warning to us all: when the same phenomena (c.f observations from a field trial) can have alternative explanations, it will require careful experimental design (and not simply more field trails) to determine what mechanisms led to that outcome. Knowing what the explanation truly is is also not just pedantic, or an indulgence for theory, for policy and actions based on an incorrect interpretation could have highly unintended consequences.
ii) C (and N) sequestration following shifts between high and low N inputs, using dairy or meat
Overall the two sets of graphs below show how the amount of C sequestered, and the rate of carbon sequestration, change over time, starting either from low N input ‘meat’ ‘M60’ (figs (a) and (b); or from higher N input dairy ‘L300’ (figs (c) and (d).
From the graphs one can see the effects of a change in fertiliser inputs per se; the effects of the shift between animal class per se; and the interactions between these. The solid lines denote where the system has become ‘meat’; the dashed lines denote where the system has become ‘dairy’. The increasing intensity of the colours (blues for dairy; reds for meat) helps spot where more fertiliser is being applied.
(Hence: ‘M60L300’ is a shift from low input meat to high N input dairy. And ‘L300M60’ is a shift from high N input ‘dairy’ to low N input ‘meat’)
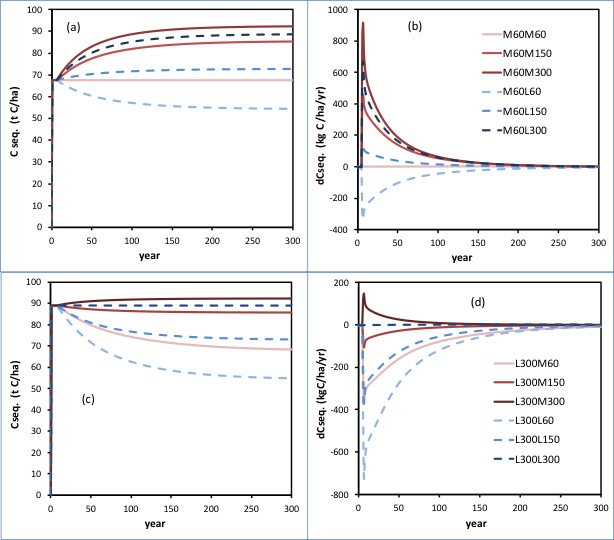
In graphs (a) and (b) it is clear how, starting from a low N input rate system, the addition of fertiliser N (greater N inputs) would be expected to increase the amount of carbon sequestered, whether the system remained a ‘meat’ system, or shifted to ‘dairy’ EXCEPT where the shift to dairy, and the consequent greater removal of N in animal products, was NOT met by the necessary increase in the rate of N inputs. Hence note the amount of sequestered carbon that could be sustained fell to a lower level in M60L60 (as we saw, and explained, in the section above).
In graphs (c) and (d) it is clear how a ‘de-intensification’ from moderately high N input ‘dairy’ systems, (‘L300’) to lower N input meat systems (‘M150’ or ‘M60’) is associated with a loss in the amount of carbon sequestered that can be sustained (hence the negative rates of C sequestration). Likewise a ‘de-intensification’ from high input dairy, (‘L300’) to lower N input dairy(‘L150’ or ‘L60’), leads to a substantial loss of sequestered carbon. This loss is considerable (c. 30 t C/ha in total) and initially at rates of C loss of c. 500kg C/ha/year).
The patterns of changes in the amounts and rates of C sequestration raise serious concerns over the capacity to understand what the final outcomes would be, from empirical studies looking (inevitably) at only the first 5 to ten years.
Theoretical (dynamic, process-based) modelling is no substitute for field observation, or even critical experimentation, but it does offer insights into what might arise long-term, on a time scale fitting for phenomena such as C sequestration, and for offering explanations for how changes in c sequestration came about. These insights can form the basis of better targeted experimentation, and not least, policy.