If you thought that cave wētā (Rhaphidophoridae) were only to be found inside New Zealand caves – Think again. They don’t need caves and they are found all around the world! A new study published in Royal Society Open Science discovered that five geologically young subantarctic islands each have their own amazing and “ancient” endemic cave wētā. [The species themselves are not ancient but each one represents a phylogenetically distinct lineage].
Pachyrhamma longicauda subadult at night on the forest floor.
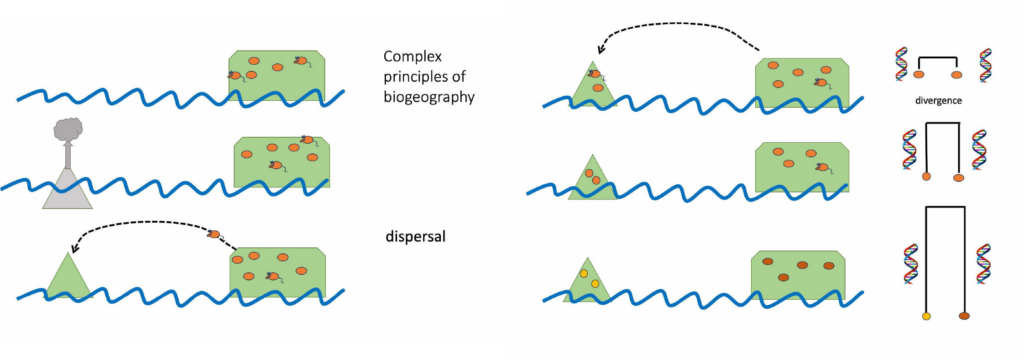
When land emerges from the sea it provides a new home for terrestrial plants and animals. Drifting across the ocean and blowing through the air new organisms arrive. New populations are established and over many generations these populations adapt to their home and might even diversify into a set of new endemic species.
Time series showing organisms colonizing new land and diverging from source population
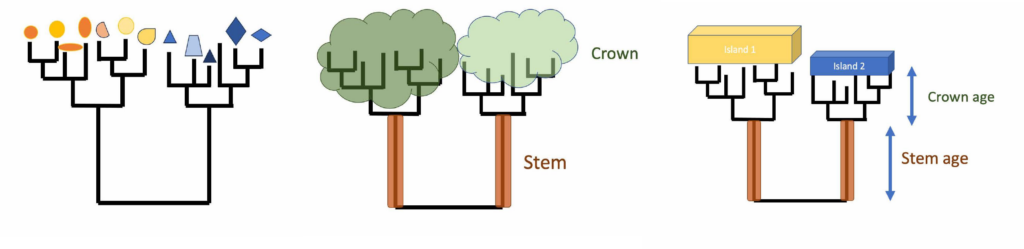
Biologists studying island endemics have long been interested in when it was that islands were first colonised after their formation, and how long it took for novel, distinguishing traits to evolve. A new study of camel crickets on seven subantarctic islands reveals how little we can learn about the time of arrival from the age of the stem. Only the age of the crown group should be used to estimate the speed of evolution.
Phylogenetic tree showing evolutionary relationships, and explanation of the terms stem and crown
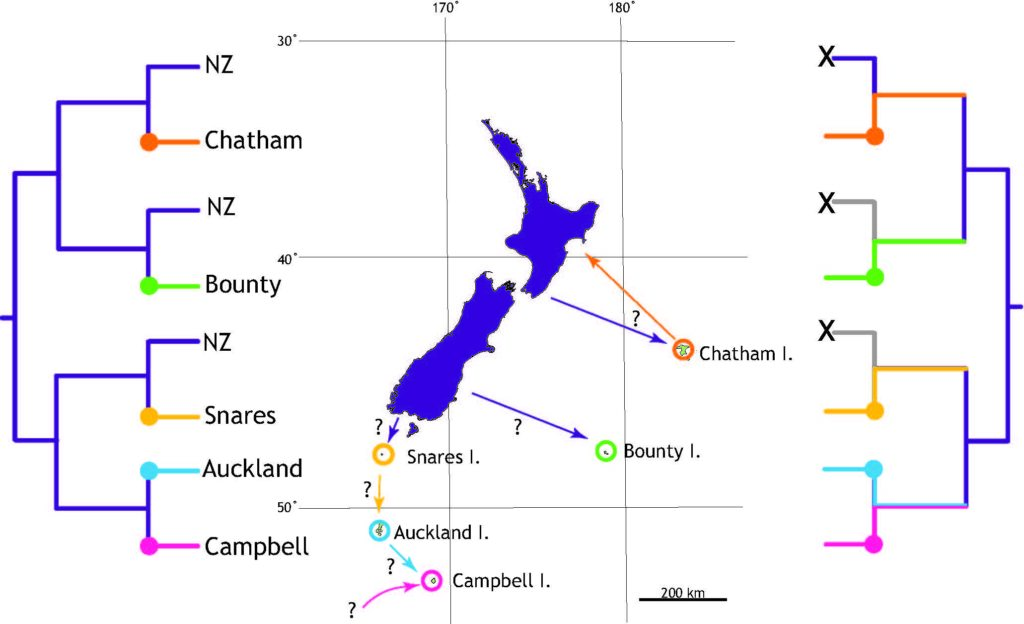
The study of Rhaphidophoridae (cave weta/camel crickets) sequenced whole mitochondrial genomes and used fossils from a sister group of orthoptera to calibrate a molecular clock. Species found only on subantarctic islands have sister taxa in New Zealand but the age of the divergence (stem) was found to be much older than the age of their island homes. In all seven cases the island species shared its most recent common ancestor with a sampled relative more than 10 million years ago (most were >25 MYA). This study provides an excellent example of how stem age can mislead biologists into thinking lineages are very old. The reason the stem dates are older than the islands is because close relatives were not sampled. All extant New Zealand genera (with one exception) were included in the study so the lack of close relatives suggests either lack of investment into species discovery or due to extinction. There is still time for us to go looking for close relatives of these island endemics in New Zealand but the islands will be smaller as sea level rises.
the tree on the right shows how extinction has resulted in very distinctive island taxa
Cave crickets are found all over the world including on tiny subantarctic islands. Finding old lineages on young islands shows that these wingless insects are successful at crossing the ocean and colonising new habitat. However, the absence of closely related species elsewhere suggests that extinction is a biologically influential factor with potential to confound traditional biogeographic assumptions
New phylogenetic analysis supports inclusion of all of the ‘alpine’ māwhitiwhiti Aotearoa in a single genus (Trewick et al. 2023).
Despite evidence of a deep ancestry of this endemic New Zealand species radiation (Koot et al. 2020) predating formation of most mountains, there is no consistent support for subdivision of the ~13 morphologically distinct species into four genera. One is sufficient and as a result the name Sigaus (Hutton 1897) now applies to all of the species.
The process of taxonomic synonymy requires care and attention to the rules set out by the International Commission on Zoological Nomenclature. The ICZN “acts as adviser and arbiter for the zoological community by generating and disseminating information on the correct use of the scientific names of animals”, and this is essential given the scale of the taxonomic challenge.
Taxonomic revision involving synonymy invariably results in binomials consisting of new combinations of genus and species epithets. This process can present a number of challenges when we reconcile names originally construed in a way that is incompatible with the current taxonomic rules. Quite often authors have not provided all the information that is required to make easy judgements and so some discussion commonly develops.
In the case of Sigaus which we infer to have been derived by Captain Frederick Hutton from the Greek σιγάω (sigáo) meaning to be silent or to keep silent. Latinisation of this word results in Sigaus and in the original combination for the type species Sigaus piliferus it is apparent that Hutton (1897) had decided on treating Sigaus as a masculine. That is useful information when formulating new combinations as is the case with these grasshoppers that including several species assigned to other genera.
If, the species epithet is adjectival then its ending may need to be altered to satisfy the Code’s requirement for gender of the binomials to match, subject to some variations. However, this does not apply if the species epithet was originally intended as a noun.
The genus Alpinacris was established by Biglow 1967 with two species: A. crassicauda and A. tumidicauda. These names, and in particular crassicauda, are used fairly frequently in zoological taxonomy, and most will immediately spot the meaning relates to thick and tail. During the establishment of new combinations the intent of the original author, Robert Bigelow 1967 in this case, can be very helpful. Was Robert coining the name crassicauda as an adjective (thick-tailed) or was he treating it as a noun in apposition (thicktail)? We don’t know because he did not record this in his description, however, the Code comes to our aid. If, the species epithet is an adjective it will need to match in gender with the genus Sigaus (masculine) and so the ending would need to be tweaked giving the combination Sigaus crassicaudus,
but if the epithet is a noun in juxtaposition the ending does not change.
The Code advises us that when the original author was not explicit then we should assume the epithet is a noun in apposition and leave it be. Hence: Sigaus crassicauda in Trewick et al. 2023.
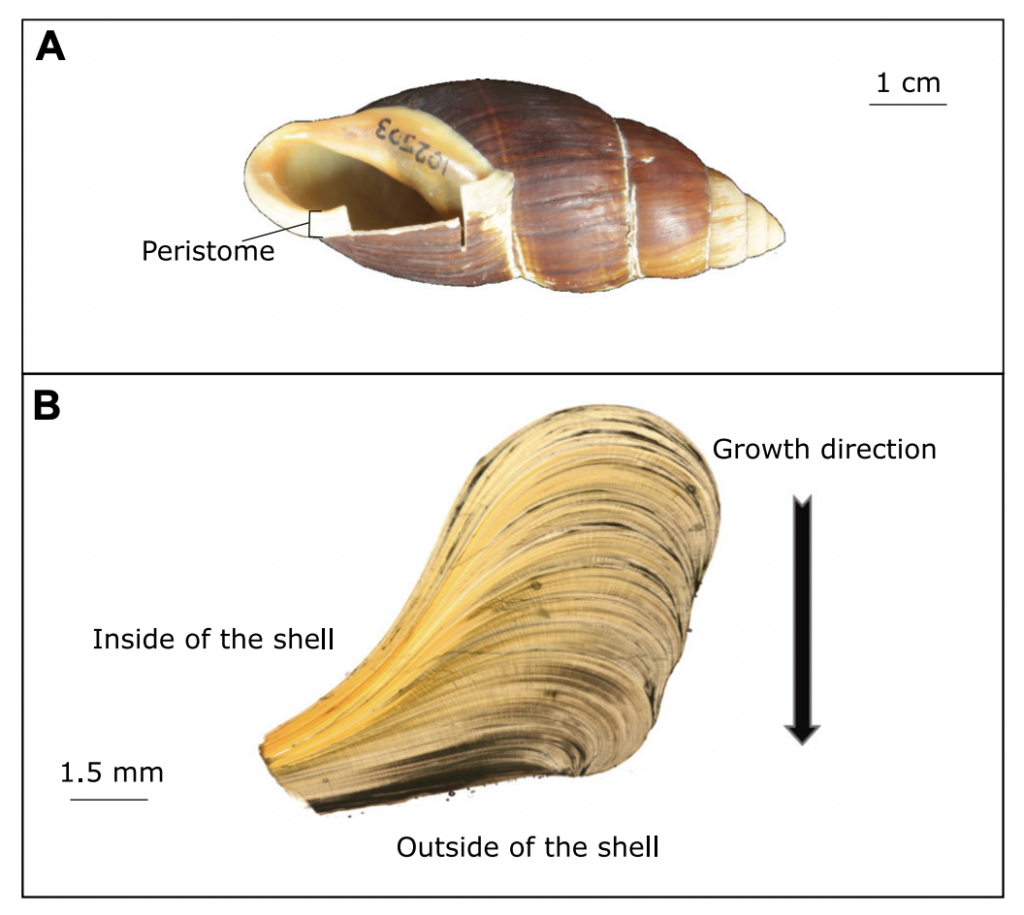
Fossil snails might tell us of the frequency of heavy rainfall in the past
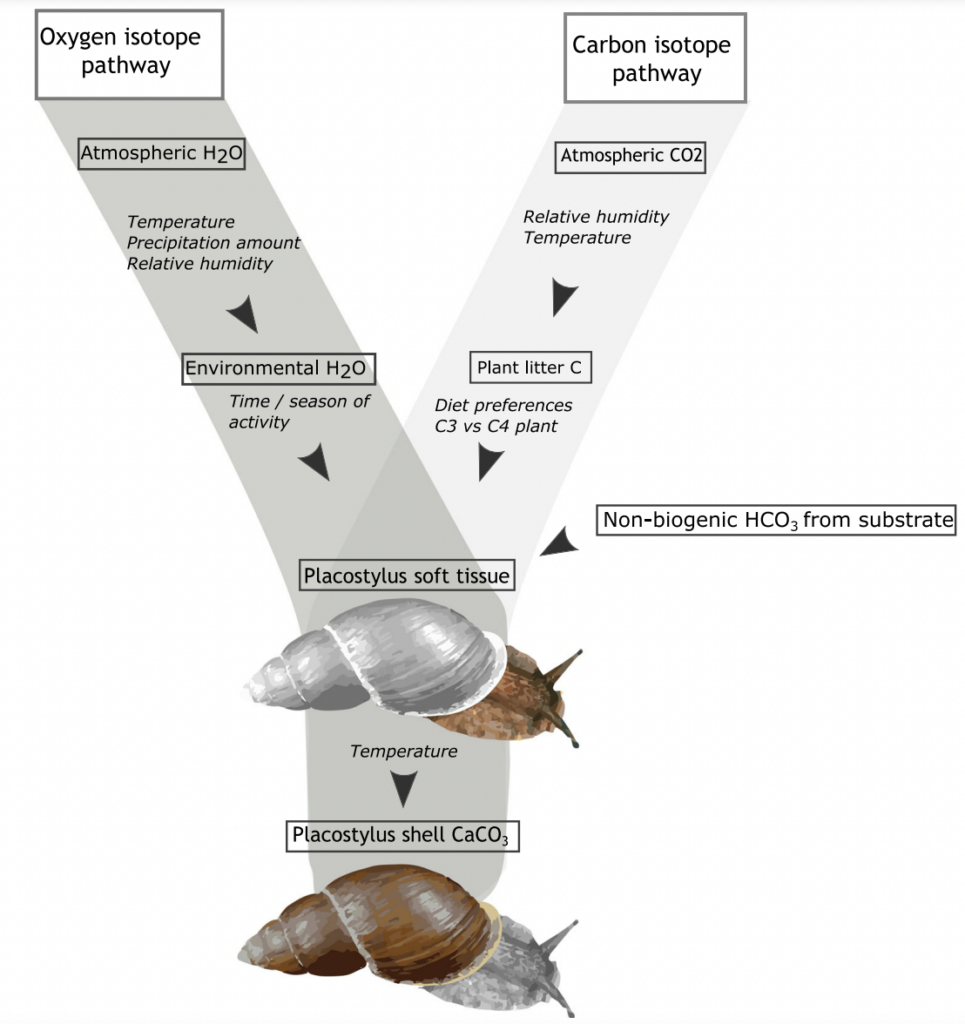
Anew study comparing the stable oxygen and carbon isotope ratios of giant land snails in New Zealand and New Caledonia found a surprising result. New Zealand snails had, on average, higher oxygen isotope ratios values than their counterparts in New Caledonia, counter to the relative isotopic composition of rainwater between these two regions. This research just published in the Journal of Quaternary Science provides baseline data for using the shells of Placostylus snails as environmental proxies – allowing us to use fossil shells to estimate the temperature and rainfall when the snails were alive.
High‐resolution stable isotope profiles from shells of the land snail Placostylus reveal contrasting patterns between snails originating from New Zealand and New Caledonia
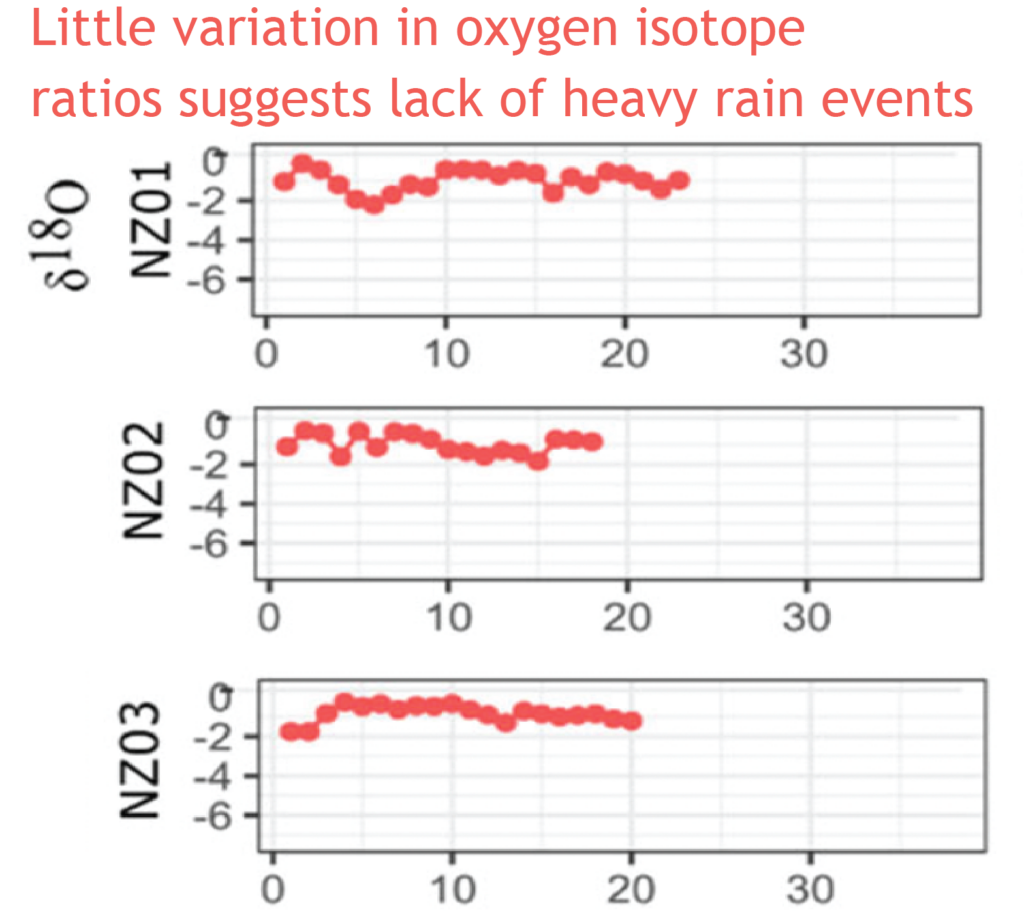
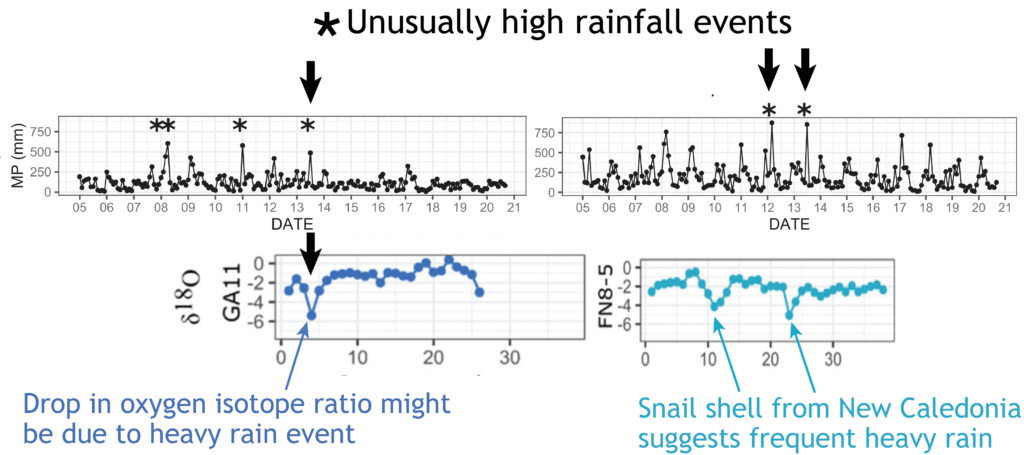
Most interesting are the dramatic drops in oxygen isotope ratios that seem to correspond to heavy rainfall – suggesting an opportunity to dig into the past to compare past precipitation with current frequency of wet weather events in New Zealand. There is also the potential to study the frequency of droughts from the pattern of snail shell growth.
Scientists at Massey University and NIWA sliced up shells of three species of giant land snail (pūpū whakarongotaua; Placostylus). The recent samples from New Caledonia showed drops in isotopic values in their high‐resolution profiles probably linked to periods of intense rainfall.
Very heavy rainfall events produce lower stable oxygen isotope ratios incorporated into the shells of the living-growing land snails. In contrast, the snails from New Zealand varied very little, suggesting that when they were alive, 74 years ago, there were few heavy rain events in the Far North of New Zealand.
The snails (pūpū whakarongotaua; Placostylus) are taonga of Ngāti Kurī who value them as security alarms (the snail that listens for war parties). Ngāti Kurī are working to save the local species from extinction but they are also kaitiaki (guardians) of fossil shells buried in the sand dunes and stored in museums. These fossil deposits could provide information about the past climate through high‐resolution stable oxygen isotope profiles.
“it is exciting to think of all the information locked up in snail shells – the shape of the shell, the DNA and the isotopes can all tell a story about the past” said Mary Morgan-Richards. “As Placostylus snails are slow growing, taking 10 years to reach maturity, and live for a long time, they can each tell their own story. There is much to be learnt by digging into old shells to reveal the frequency of heavy rainfall events in the past.”
To get ahead in science I recommend you take a distinctive name
Any one who publishes will know that they want their name to stand out among the crowd and be unique to them. If you set up an email account or use twitter you have probably invented a name for yourself.
I changed my name when I started publishing science articles when I was about 27 years old. Up until then I had three names: first, middle and last (surname). My second (middle) name was the same middle name as my two sibblings, my father and a couple of cousins. The name came from my great-grandfather who was Rev. Morgan Richards. He married a girl with the surname Morgan and they then used it as a middle name for their two sons. The elder, my grandfather, and his elder son carried on the tradition. Providing me with a middle name that was also a ‘family’ name.
There are many reasons why it helps to have an unusual name. For a career in science it does help to be easily found and cited. You don’t want your views confused with the crazy professor and you do want to be contacted for the opportunity to give a keynote speech at an international conference in Madeira. Try searching the world-wide-web and see how many people share your name. When I narrowed my search down just to people who write in science journals, my double-barrelled last name was unique.
Searching using the author tool in the “scopus” database finds many people (authors) with the same name
When I was looking back over 25-years of Molecular Ecology meetings in NZ I tried to find where the original participants had moved to. Some people were easy to track down just because they had an interesting combination of initals, but many names were surpringly common. I never did find the Luis Medrano, Alex Quinn or James Bower I was looking for.
Hyphen or not?
Initally I thought I’d emulate John Maynard Smith and have a double surname without a hyphen – but it was hard to get other people to see the difference between John Maynard Smith and John Maynard Smith. Try using a software tool for formating and inserting references and you’ll see that the designers didn’t consider recognising Maynard Smith as a double barrelled surname. So, I add the hyphen. 20 years later I had a student who tried to be B. Taylor Smith without a hyphen and she pullled it off for her first paper but then gave up and converted to Dr Taylor-Smith.
Over the years I became Dr Morgan-Richards and have never regetted it. I do feel its a pity both names come from my fathers side, but lots of people take their partner’s father’s name – so at least I’m related to both originals. My mother’s father’s name is ‘Wild’ which might have been more fun to play with but would I have choosen to be so close to the end of the alphabet? There are still plenty of lists when the Abbots come long before the Witches. And you don’t want to queue for research funding with a name at the end of the list.
Family names
Some people like the idea of sharing a surname – even one that is quite boring. And if the guy is ready to change his name to her name then that feels like an imbalance is being redressed. At the school gate you’ll be able to match child with parent using just their last name but I think there must be better ways to do that. Once you are famous you’ll probably want to keep your own name but creating a new double barrel surname that you both use is a great solution if the names are short – I think the kids of Dr Charlesworth and Dr Pattabiraman might not thank their parents for creating a unique mouthful of a last name. One of my good friends gave her first born child her husband’s surname, and gave her second child her own surname. That way each offspring had one parent easily identifed on letters to the school (e.g. please excuse little Moana from class today…. signed person with the same surname (must be mother/father)).
I could have formally changed my name at no cost when I got married in 2000 – don’t know why I didn’t. I blame baby-brain. The only time it mattered was getting airline tickets in the same name as that on my passport – and that really does matter. So, I paid to changed my name some years later. Now its offical and I’ve been using this name for almost 30 years- but that doesn’t change how my mother and sister and sister-in-law address their letters to me. You can put an interesting name on a girl but you can’t change her family.
My family and friends have different names for me but the message is the same
What controls the colour of the common mānuka stick insect?
Three stick insects . The same species but different colours living in different environments
When we see how well-camouflaged individual stick insects are, it is hard not to imagine they have chosen either their hiding place or their skin colour, with care. But stick insects cannot see colour – they don’t know if they are green or brown. The reason they are so hard to see is that their cousins who were visible have been eaten. The only ones left are the ones that were not seen by the warblers and silver eyes and chaffinches and tui and sparrows.
Some stick insects can change colour – as demonstrated by the Indian stick insect Carausius morosus (Mangelsdorf 1926), whose colour changes in response to light levels. The trick this insect uses is the movement of pigment granules in it’s skin (cuticle) that alter light absorption and light scattering (Umbers et al. 2014).
Other stick insects like the North American Timema walking sticks have their colour determined by their genes and variation among individuals within the same species is maintained by selection, recombination and mutation (Comeault et al. 2015; Villoutreix et al. 2020). They cannot change colour to suit their environment.
So, what about rō Aotearoa – New Zealand stick insects? No one knows for sure – but maybe you have an idea? I have made some observations of the common mānuka stick insect Clitarchus hookeri and think that genetics is involved.
Colour variation in captivity suggests genetic rather than environmental differences explain this phenotypic diversity
Nymphs of the mānuka stick insect Clitarchus hookeri showing the range of colours a single individual can exhibit at different instars
In boxes at home I allowed green Clitarchus hookeri indivudals from Waikato to mate with brown individuals from Whanganui. I collected the eggs that each female dropped and stored them for a few months until the little stick insects hatched. Nymphs were housed in sibling groups and fed mānuka (Leptospermum scoparium) and climbing rātā (Metrosideros perforate).
All my Clitarchus hookeri offspring hatched as green nymphs. Many remained green as they grew and moulted but some changed colour (see figure). Two males changed to brown when about 30mm long, then speckled grey, before moulting to green as adults, their sisters remained green as they grew. Two females changed to brown when about 25mm long, then pale brown. One matured as a green individual, the other pale brown. Of the twelve stick insects I raised until they were adult eight remained green and four changed colour but only one was still brown when adult. In the next generation, produced by crossing siblings, the similar observations were made; some F2 individuals remained green as they grew, some changed to brown when about 30mm before changing to green as adults.
My observations of the colour of F1 offspring provide preliminary data suggesting that the colour of C. hookeri individuals is genetically determined rather than environmental as nymphs were raised in a common environment. However, I also observed that individuals can be different colours at different stages of their growth. Although changing from green to brown has been recorded in this species (Stringer 1970), changing back to green from brown was unexpected. Individual variation in colour will need to be considered when modelling inheritance and selection of this trait. It is possible that my observations could be explained by relatively few alleles at one or two loci. Further work is needed for us to understand the inheritance of colour traits in the common mānuka stick insect Clitarchus hookeri.
References
Comeault AA, Flaxman SM, Riesch R, Curran E, Soria-Carrasco V, Compert Z, Farkas TE, Muschick M, Parchman TL, Schwander T, Slate J, Nosil P. 2015. Selection on a genetic polymorphism counteracts ecological speciation in a stick insect. Current Biology. 25:1975–1981.
Mangelsdorf AJ. 1926. Color and sex in the Indian walking stick, Dixippus morosus. Psyche. 33:151–155.
Morgan-Richards M, Trewick SA, Stringer IAN. 2010. Geographic parthenogenesis and the common tea-tree stick insect of New Zealand. Molecular Ecology. 19:1227–1238.
Morgan-Richards M, Langton-Myers SS, Trewick SA. 2019. Loss and gain of sexual reproduction in the same stick insect. Molecular Ecology. 28:3929–3941. DOI: 10.1111/mec.15203.
Salmon JT. 1991. The stick insects of New Zealand. Auckland, New Zealand: Reed.
Stringer IAN. 1970. The nymphal and imaginal stages of the bisexual stick insect Clitarchus hookeri (Phasmidae: Phasminae). New Zealand Entomologist. 4:85–95.
Trewick SA, Morgan-Richards M. 2005. New Zealand Wild: Stick Insects. Reed Publishing: Hong Kong.
Umbers KDL, Fabricant SA, Gawryszewski FM, Seago AE, Herberstein ME. 2014. Reversible colour change in Arthropods. Biological Reviews. 89:820–848.
Villoutreix R, de Carvalho CF, Soria-Carrasco V, Lindtke D, De-la-Mora M, Muschick M, Feder JL, Parchman TL, Gompert Z, Nosil P. 2020. Large-scale mutation in the evolution of a gene complex for cryptic coloration. Science 369:460–466.
Sweet chestnuts are falling from the tree in my garden (late March in Manawatū, NZ). We collect them all up so the rats, rabbits and rosellas don’t get them. I simmer whole nuts in a large pot of water for about 15mins the same day I collect them. Once cooked they can be easily opened with a sharp knife. Cut them open while still quite hot and scoup out the soft cream flesh. This nutty pulp can be frozen and used later in the winter for making stuffing for a roast chicken, thickening soup, or used in baking.
One option is to use chestnut pulp instead of ground almonds when baking. For example, twice-baked biscuits, dry and crunchy are usually made with almonds but work well with chestnuts. Biscotti is not super sweet but perfect for dipping into a morning coffee or extra fuel before the cycle home. I take a screw-top jar filled with biscotti to work and they prevent me seeking chocolate when I’m busy. So, here’s a recipe for sweet chestnut biscotti to try when the nuts are falling:
Ginger and sweet chestnut biscotti 25 fresh chestnuts (makes about 250g pulp) 3 small eggs or 2 large (supermarket-type) eggs 1tsp vanilla essence 1/4tsp salt 100g sugar (I used half white, half brown) 120g crystalised ginger 70g cranberries (or other dried fruit) 1.5 – 2 cups plain white flour 1tsp baking powder 1tsp ground ginger
Beat the eggs with essence and salt. Add chestnuts, crystalised ginger, dried fruit, and sugar. Stir. Add flour, baking powder and ground ginger then mix well to form a dough that is dry enough to turn out and shape with your hands. Kneed the mixture on a floured bench briefly then shape into a log about 30cm long. Move log to oven tray and bake for 40–50 mins at 160oC. Remove from oven and while warm cut with a bread knife into slices about 1cm thick. Arrange these on an oven tray and return to the oven. Bake about 15 mins then turn every slice over and bake another 15 mins. The exact time will depend on your oven and how crunchy you like your biscotti. Turn oven off and leave to cool in the oven. Store in an air-tight container.
If you don’t like ginger, you could try swapping spices and increasing dried fruit. For example here’s a version for cinnamon, apricot and chestnut biscotti (mix and bake as above)
50 fresh chestnuts (makes about 500g pulp)
6 small eggs or 4 large (supermarket-type) eggs
1tsp Vanilla essence
zest from one lemon
200g sugar
2.5 cups plain white flour
15 dried apricots (or other dried fruit)
1tsp baking powder
1tsp ground cardamon
1tsp ground cinnamon
1/2tsp salt
Parma Tarts – Medieval chestnut and chicken pies
600g chicken mince
Oil for frying (couple of tablespoons canola)
500g chestnut pulp (about 50 chestnuts)
½ cup currants
1 tsp freshly ground black pepper
1 tsp ground ginger
1tsp cinnamon
½ tsp mixed spice
1tsp salt
2 small eggs (or one large egg)
Fry chicken mince in lots of oil until colour change and looking cooked. Add currants and spices. Mix well then add chestnuts. Keep frying and stiring for a few more minutes. Turn heat off and add one or two eggs and mix well to bind.Line pie tins with flakey pastry and fill with meat. Top with pastry lid and bake at 180oC until pastery golden. Yum!
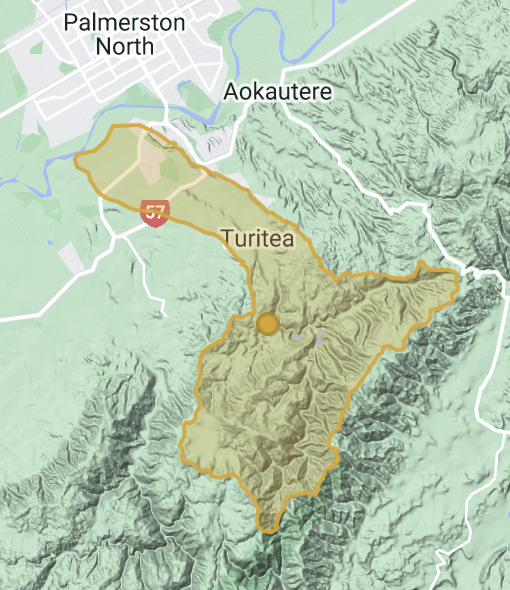
TURITEA VALLEY is like many in Aotearoa New Zealand. A water catchment cut by a small water course (a tributary of the Manawatu River), that was formerly shrouded in dense native forest. Today the stream and its catchment is highly modified with little of the former native vegetation, extensive pasture development and increasing residential development. This project seeks to collate information about native and introduced biota as we develop our understanding of the way people interact with natural ecosystems. It should be possible for us to maintain native biodiversity, water quality, land stability and ecosystem processes at the same time as drawing on the natural resources our landscape offers.
One tool we use is iNaturalist, which is “is a social network of naturalists, citizen scientists, and biologists built on the concept of mapping and sharing observations of biodiversity across the globe”. iNaturalist NZ—Mātaki Taiao is our local rendering of that global resource and it is easy to get involved if your start here.
Within iNaturalist NZ we have set up a project called Turitea, which allows us to easily view and organise observations made by iNaturalists within the Turitea catchment. It is situated at the north west end of the Tararua range just south of the Manawatu Gorge, draining low hills and flowing west through the Manawatū campus of Massey University to the Manawatu river. The headwaters of the Turitea stream provides drinking water to Palmerston North city. Any observation made within the bounds of the water catchment can be analysed.
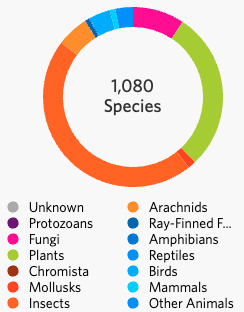
As of 17 March 2023 the Turitea project on iNaturalist had accumulated 3,507 observations spanning 1,080 species. Some species are documented many times and that is useful because it can show activity across the landscape and through time. For example the native pigeon kereru was observed most often in Turitea during September and a large proportion of sightings were in residential areas and on Massey University campus.
Large species are most often spotted and recorded (above) but the range of organism types is huge and includes native and introduced species (below).
There tends to be a bias towards animals but plants, algae and fungi are fundamental to the ecosystem. Ninetynine species of fungus and lichen have been recorded in 264 observations, suggesting that any additional observation is likely to contribute new species to the list. Most fungi are growing under or on trees and this reflects their importance in nutrient cycling in forests and their relative scarcity in exotic pasture grasslands. ‘Mushroom’ type fungi (59 species) are the most readily spotted and recognised but fungi are also present in encrusting lichens (6 species), brackets (5 species) and as insect parasites.
Gilled mushrooms in Turitea bush
Some fungi operate as parasites and these include species of Cordyceps which invade insect hosts and gradually take over the hosts body. Cordyceps sinclairii (left) lives on the late stage nymphs of native cicadas, and produces a small fruiting body with many white spores in Autumn. Other species have ground living moth larvae as their hosts and some are parasites on adult insects. The infamous ‘vegetable caterpillar‘ has not yet been recorded in Turitea but might exist under the more intact forest of the water reserve.
The distribution of plant species depends a lot on landuse practices. Areas modified for pasture are dominated by a few exotic grass and herb species, whereas remnant native forest have a richer species mix. Notable native trees include northern rātā which is represented in the valley by a small number of individual trees, kamahi, and rewarewa. Ferns include bracken in relatively undisturbed open areas, and many species in remnant forest.
Pest species abound in the Turitea valley and among mammals include the usual suspects: ferret, rat, rabbit possum (pictured), hedgehog, stoat, mouse, feral cat. However only 40 observations have been recorded suggesting that other ways are needed to get a better understanding of pest abundance. Small scale trapping schemes that are regularly monitored probably have limited impact on total pest numbers but do provide invaluable data about the abundance of species targeted.
Monitoring changes in species composition, abundance of particular species and changes in activity patterns over time (seasons or years in the face of climate change) is best gained by repeated observations at a particular sampling site. This can be done at any scale convenient to the observer such as recording night flying moths arriving at a house window. So far 154 species of moth and butterfly have been documented in Turitea valley (666 observations), but one site (554 observation) includes 128 species
And observations don’t have to be visual. The natural sounds in the valley can get drowned out by cars and lawnmowers but they are there to heard. These include katydid, cicada, introduced whistling frog, ruru more pork, and whitehead which are becoming more frequent in parts of the valley.
Only two native non-bird vertebrates are known from the valley and from just one location so far. Both are reptiles: a Naultinus green gecko is know from a single dead individual. A small population of New Zealand grass skink (left) is known, but others probably exist… An invasive skink has also been detected.
At the other end of the spectrum, one of the smallest (about a fifth of a millimetre long) and hardiest of organisms so far recorded in the valley is a kind of tardigrade or ‘water bear’ that belong to their own phylum Tardigrada.
Global climate change is changing everything, but the gradual processes make it hard to spot the extent of the impact. Change in local climate and the effects it has on species and ecosystems are most apparent where there is a steep gradient in conditions.
A good example of a steep environmental gradient can be found on any tidal rocky coastline, where the tide means some creatures live under sea water, others are exposed to air occasionally, while others that live further up the beach are exposed to drying for much longer. All parts are physically near each other making the gradient in conditions steep. Coastal environments are impacted by global climate change because warming results in the melting of glaciers and polar ice, which in turn leads to rising sea level.
Tree-line on Mount Arthur, Nelson Lakes New Zealand.
On land, a similar situation exists on mountains because the slope of the mountainside means nearby places can have very different conditions. Most obvious as you move up a mountain is the lowering temperature. It is colder near the top than the bottom which is why you go up a mountain to find a ski field.
Another feature of mountains that has a strong influence on biological diversity is their tendency to form ‘islands’; patches of alpine habitat in a sea of lower elevation conditions which in New Zealand is normally forest. Valleys, rivers and forest create a patchwork of mountain tops and ridges; connectedness of these habitats and depends on the climate gradient.
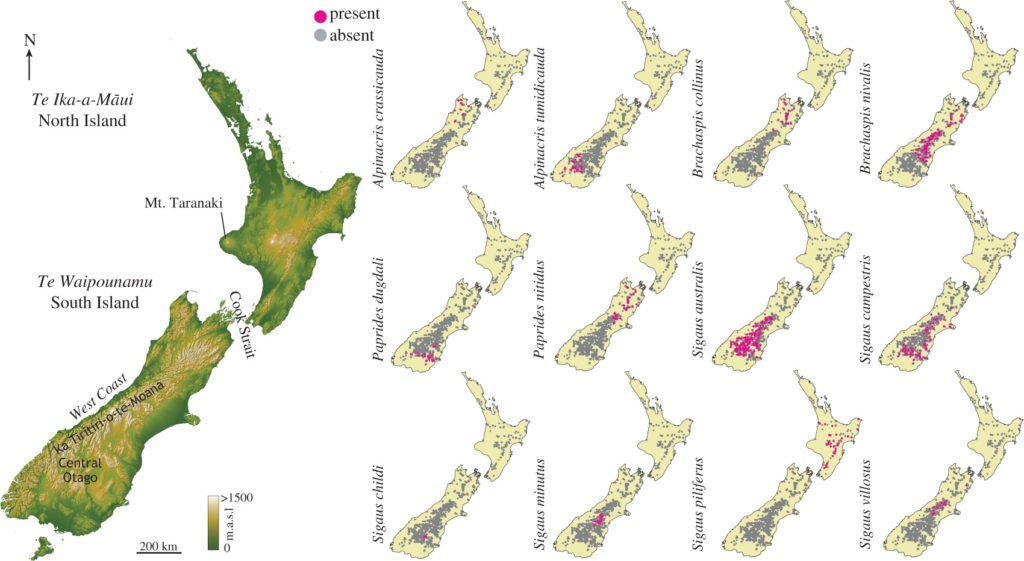
Animals, plants, microbes and fungi that live in the alpine zone, such as this grasshopper (Sigaus piliferus on Mount Ruapehu) have evolved to thrive in the conditions of extreme day-night and seasonal fluctuations in temperature and water availability. Survival means coping with all the different conditions, including being able to freeze when it is cold and re-animate when it is warmer.
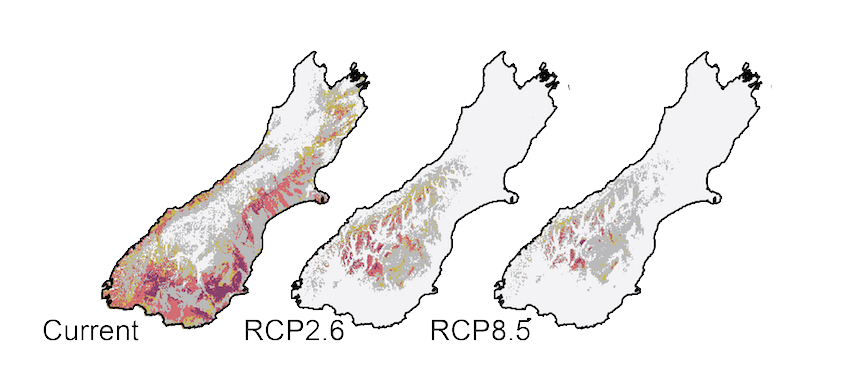
Locations of presence and absence for each of 12 New Zealand grasshopper species
Using the known distribution of New Zealand’s endemic alpine grasshopper species we identified their preferred habitat based on climatic conditions, and then modelled the future status of those habitats given anthropogenic climate warming. We found that available habitat will change for these alpine creatures very quickly; in about 70 years some species could be without suitable habitat that they can use.
It is easy to measure environmental conditions such as average temperature today, and good data about conditions in the past come fromice-cores and other sources, the future is more uncertain. We know the physics that connects atmospheric gases with global temperature, but the future depends on what people do. We can predict temperature changes during the rest of this century forseveral scenarios which are based upon the balance between the Earth’s heat (energy) gain and its loss (radiative forcing). The most extreme scenario used by the IPCC, RCP8.5, would result with from continued increase in GreenHouse Gases emissions. A more optimistic, but highly unlikely scenario given current trends, RCP2.6, would involve reduction in GHG emissions starting in 2010. RCP2.6 will still result in a 2˚ mean global temperature rise by 2100 (compared to 1750), compared to about 5˚ under RCP8.5.
Current and predicted available habitat for the endemic New Zealand grasshopper Sigaus australis (dark colours most suitable). The RCPB.5 climate change scenario assumes GHG emissions continue to rise through the 21st century. Even under the optimistic RCP2.6 that assumes the C02 emissions started declining in 2020 (they did not) and continue to O by 2100, habitat for S. australis will be scarce. RCP2.6 would result in a global average temperature increase of about 2˚ in the next 70 years.
Our findings apply to all biology living in the alpine zone and, by extrapolation, to all biology in New Zealand. Within one human lifetime, habitat availability will have changed catastrophically for many species… Others may gain, but these will often be species that humans have moved from their native habitat, and become weeds and pests.
Pūpū whakarongotau (Placostylus ambagiosus) is a large leaf-eating land snail that has declined to fewer than 2000 individuals scattered over 19 populations. These snails are highly valued by tangata whenua of far north Aotearoa (Te Aupōuri me Ngāti Kurī), because in the past the snail was both kai and made alarm calls at night warning of approaching invaders. The sounds these pūpū (snails) make as they hastily retreat into their shells when disturbed at night once alerted the people to approaching invaders and so saved their lives. So, the snails are known as pūpū whakarongotaua -the snail that listens for the war party. Oral histories tell us that snails were moved to propagate new P. ambagiosus populations along with harakeke and karaka.
We know that individuals of this species seldom move more than a few metres from where they hatch, are long-lived (10–22 years), and show strong site fidelity – with individual snails being able to crawl home over at least 60 metres (Parrish et al. 2014; Stringer et al. 2017). The tough shell protects adult snails from native predators and the climate, and preserves evidence from the past.
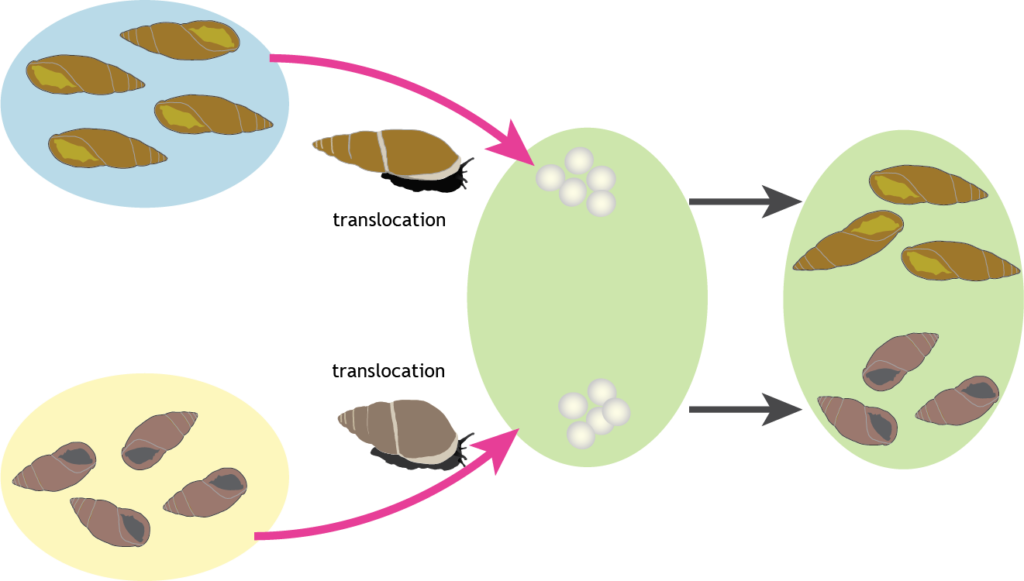
Visible differences in the size and shape of snail shells of this species (Placostylus ambagiosus) led to numerous distinct isolated populations being given their own subspecies name. Using museum material collected 70 years ago we studied shell shape variation to determine whether it is the result of genetic differences or environmental differences. On a headland, previously the site of a pā (fortified settlement), one population that resulted from prehistoric cultivation of snails showed that shell shape differences are maintained when the snails are living and growing in the same environment.
A “common garden” experiment. Snails moved from different populations to the same area continue to show their distinct shell shape characteristics over generations, revealing their genetic differences.
Using geometric morphometrics of shell shape we could discriminate pūpū shells without reference to where they had been collected. Our genetic data confirmed that some human movement of snails had occurred but that this has not resulted in a loss of genetic differentiation from east to west. We recommend that the shell shape (not size) of these species can be used to infer genetic differences that might be important for the survival of the species as climate changes. All shape variation should be conserved by protecting all living populations from predators and competitors. The subspecies names are a good way to refer to this diversity and protect the evolutionary potential and historic record held in these populations.
View from Mount Pitt to the Norfolk Island airstrip and Phillip Island beyond.
In the Pacific Ocean, east of Australia and about halfway between New Zealand and New Caledonia is Norfolk Island. This tiny island (~30 square kilometres) has a colourful history and enough endemic species to make it very valuable to biologists. Not as famous as the Galapagos Islands or the Hawaiian archipelago, Norfolk Island has its own endemic biodiversity including dozens of species of tiny snails in the forest leaf litter.
During WWII an aeroplane runway was constructed for refuelling Royal New Zealand Air Force (RNZAF) bomber patrols and for a transport service to Bougainville. Construction required relocating families and cutting down Norfolk pines (Araucaria heterophylla), and the runway takes up a huge portion of the land surface of Norfolk Island . The giant ‘T’ of 2.22km and 1.8km imprinted across an island that is only a little over 9km at its widest point. Access by commercial airlines now brings tourists and invasive species to the island on a regular basis. It only takes a couple of hours flying from Auckland to reach Norfolk Island, and there are flights in and out a couple of times each week. Before the runway and the ships that brought people, new species to the island arrived by long-distance dispersal – flying, swimming, ballooning, rafting, and hitch-hitching (Jordano 2016). Consider the many birds that fly to Norfolk Island, some are capable of bringing live snails in their guts. For example, some snails can survive being swallowed by silver-eyes and be found alive in their droppings. These little birds are found on many oceanic islands so can aid dispersal of snails (Zosterops sp. Wada et al. 2012). Ducks (also seen on Norfolk Island) can transport some snail species on the inside (Van Leeuwan et al. 2012). Seabirds, of which Norfolk Island has eight breeding species, are also potential source of long-distant dispersal (Viana 2016).
A Norfolk Island robin, part of a species complex on Pacific islands.
Norfolk Island was formed about 3.05–2.3 million years ago from several volcanic eruptions (Jones & McDougall, 1973). The terrestrial fauna of Norfolk Island must therefore have developed in just a few million years (<3) from the descendants of long‐distance dispersing ancestors (Holloway, 1977). Many plants and invertebrates endemic to Norfolk Island look similar to species elsewhere in the Pacific. Their ancestors must have dispersed to colonise this volcanic island. Isolated on Norfolk Island, populations have accumulated differences (allopatric speciation) and in many cases can be readily distinguished as similar but different from Australian or New Zealand species. For example, the endangered Norfolk Island coastal shrub Coprosma baueri looks very like New Zealand taupata Coprosma repens and the Norfolk Island Palm (Rhopalostylis baueri) looks like New Zealand nikau (Rhopalostylis sapida). When the Norfolk Island boobook owl (Ninox novaeseelandiae undulata) was down to a single female, it was genetically similar enough to successfully hybridise with a male ruru from New Zealand (Ninox n. novaeseelandiae; morepork) to save the population from extinction (Garnett et al. 2011). The cicada species found on Norfolk Island, Kikihia convicta, is morphologically and genetically sister to the New Zealand species K. cutora (Arensburger et al. 2004). And sister relationships between NZ and Norfolk Island taxa are also seen with the extinct kaka (parrot; Nestor productus) and extinct pigeon (Hemiphaga novaeseelandiae spadicea; Goldberg et al. 2011). These Norfolk Island species have New Zealand affinities, but many others have close relatives in Australia, New Caledonia and other Pacific Islands. For example, of the larger butterflies and moths native to Norfolk Island about 22% are endemic, of which only 10% have New Zealand origins (Holloway, 1977).
Species Radiations
Arboreal snail.
Cryptochropa exagitans
Fanulena insculpta
Allenoconcha basispiralis
Roybellia platystoma
Telmosena suteri
Greenwoodoconcha nux
Palmatina quintali
There is considerable species diversity of terrestrial micro-snails in Norfolk Island, best estimates are that there are about 40 living species (Neuweger et al. 2001; Varman 2016). Most are known only from empty shells, and species descriptions of micro-snails usually rely on shells (Stanisic et al. 2010). While on Norfolk Island we focused our effort of getting photographs of the live snails of the common species . We collected snails from the ground from forest leaf litter and from leaves during the day.
We photographed living specimens from as many common species as we could find, with the hope that this resource can be used to provide better tools for their identification in the future and help people conserved the current snail diversity. For example the Pinwheel snail Cryptocharopa exagitans (from the family: Charopidae) is recorded (from empty shells) as the most common micro-snail in mixed forest leaf litter near Duncombe Bay, Norfolk Island (Neuweger et al. 2001). The shell of this pinwheel snail (3.5mm) is recognised by its frill of dried mud around the edge but when alive the tiny snail shell is fantastically camouflaged as rock so very easily over-looked. Our photos show the tiny snail hauling what looks like a stone on its back.
Leaf Beetles Two species of eumolpine leaf beetle are described from Norfolk Island (Dematochroma shuteae and Dematochroma norfolkiana Jolivet et al. 2007), although it is likely there are more to study. The adult beetles are small and usually brown, bronze or black. They feed at night on leaves but as adults the beetles are probably short lived and likely to be seasonal. Larvae of these beetles live underground feeding on roots. The two known species are probably related to the eumolpine radiation of New Caledonian beetles (Gómez-Zurita 2011), also found in New Zealand and Australia. The leaf beetle species on Lord Howe, look quite different from one another but represent an island radiation from a single recent ancestor. We hope to find out whether the Norfolk Island leaf beetle species also represent an endemic radiation.
Norfolk Island Eumoplinae
By searching in the Norfolk Island leaf litter during the day and at night on foliage we saw numerous individuals of at least four different types/forms that might represent four species. Most beetles were active during the night when mating pairs were frequently observed. One species was predominantly observed on the foliage of Piper excelsum psittacorum, and another species was seen on leaves of Coprosma pilosa. Leaf beetles often have a short season as adults, so one week of observations is likely to have included just a fraction of all Norfolk Island Eumolpinae beetles. We are fairly confident that work on these insects will double the known diversity of leaf beetles from Norfolk Island.
References
Arensburger P, Simon C, Holsinger K. 2014. Evolution and phylogeny of the New Zealand cicada genus Kikihia Dugdale (Homoptera: Auchenorrhyncha: Cicadidae) with special reference to the origin of the Kermadec and Norfolk Islands’ species. Journal of Biogeography 31: 1769-1783. Goldberg et al. 2012. Population structure and biogeography of Hemiphaga pigeons (Aves: Columbidae) on islands in the New Zealand region. Journal of Biogeography 38: 285-298. https://doi.org/10.1111/j.1365-2699.2010.02414.x Gómez-Zurita J. 2011. Rhyparida foaensis (Jolivet, Verma & Mille, 2007), comb. n. (Coleoptera, Chrysomelidae) and implications for the colonization of New Caledonia. ZooKeys 157: 33-44. Holloway, J.D. 1977. The Lepidoptera of Norfolk Island. W. Junk, The Hague. Jolivet, Verma & Mille 2007. New species of Dematochroma from Lord Howe and Norfolk Islands (Coleoptera, Chrysomelidae, Eumolpinae). Nouv. Revue Ent. 23; 327-332. Jones, J.G. & McDougall, J. 1973. Geological history of Norfolk and Philip Islands, southwest Pacific Ocean. Journal of the Geological Society of Australia 20: 239–257. Neuweger et al. 2001. Land Snails from Norfolk Island Sites. Records of the Australian Museum Supplement Nov. 2001. DOI: 10.3853/j.0812-7387.27.2001.1346 Reid C. 2003. Chrysomelidae of Lord Howe Island. Chrysomelidae, 42: 7. Van Leeuwen et al. 2012. Experimental Quantification of Long Distance Dispersal Potential of Aquatic Snails in the Gut of Migratory Birds. PloS One. https://doi.org/10.1371/journal.pone.0032292 Varman R. 2016. Norfolk Island Snail Species Collections made between January and March 2016. Report to Australian National Parks. Viana et al. 2016. Migratory Birds as Global Dispersal Vectors. Trends in Ecology and Evolution 31: 763-775. Wada, et al. 2012 Snails can survive passage through a bird’s digestive system. Journal of Biogeography 39: 69-73